Промежуточный мозг является конечным отделом мозгового ствола и сверху полностью покрыт большими полушариями. Основными образованиями промежуточного мозга являются таламус (зрительный бугор) и гипоталамус (подбугровая область). Последний соединен с гипофизом — главной железой внутренней секреции. Вместе они составляют единую гипоталамо-гипофизарную систему.
Промежуточный мозг интегрирует сенсорные, двигательные и вегетативные реакции организма. Он подразделяется на таламус, эпиталамус и гипоталамус.
Таламус представляет своего рода ворота, через которые в кору поступает и достигает сознания основная информация об окружающем мире и о состоянии тела. Таламус состоит примерно из 40 пар ядер, которые функционально делятся на специфические, неспецифические и ассоциативные.
Специфические ядра служат областью переключения различных афферентных сигналов, направляющихся в соответствующие центры коры головного мозга. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы и внутренних органов. Эти структуры осуществляют регуляцию тактильной, температурной, болевой и вкусовой чувствительности, а также зрительных и слуховых ощущений. Так, латеральные коленчатые тела являются подкорковыми центрами зрения, а медиальные — подкорковыми центрами слуха. Нарушение функций специфических ядер приводит к выпадению конкретных видов чувствительности.
Основной функциональной единицей специфических ядер таламуса являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору больших полушарий от кожных, мышечных и других рецепторов.
Неспецифические ядра являются продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса. Неспецифические ядра таламуса диффузно посылают нервные импульсы по множеству коллатералей ко всей коре головного мозга и образуют неспецифический путь анализатора. Без этого пути информация анализатора не будет полной.
Повреждения неспецифических ядер таламуса приводят к нарушению сознания. Это свидетельствует о том, что им пульсация, поступающая по неспецифической восходящей системе таламуса, поддерживает уровень возбудимости корковых нейронов, необходимый для сохранения сознания.
Ассоциативные ядра таламуса обеспечивают связь с теменной, лобной и височными долями коры больших полушарий. Повреждение этой связи сопровождается нарушениями зрения, слуха и речи.
Через нейроны таламуса вся информация идет в кору головного мозга. Таламус выполняет роль «фильтра», отбирая наиболее значимую для организма информацию, которая поступает в кору больших полушарий.
Таламус является высшим центром болевой чувствительности. При некоторых поражениях зрительного бугра появляются мучительные болевые ощущения, повышение чувствительности к раздражителям (гиперестезия); незначительное раздражение (даже прикосновение одежды) вызывает приступ мучительной боли. В других случаях нарушение функций таламуса вызывает состояние анальгезии — снижение болевой чувствительности вплоть до полного ее исчезновения.
Эпиталамус
Эпиталамус, или надбугорье, состоит из поводка и эпифиза (шишковидная железа), которые формируют верхнюю стенку третьего желудочка.
Гипоталамус
Гипоталамус располагается вентральнее зрительного бугра и является главным центром вегетативных, соматических и эндокринных функций. В нем различают 48 пар ядер: преоптические, супраоптическое и паравентрикулярное, средние, наружные, задние. Большинство авторов выделяют в гипоталамусе три основные группы ядер:
передняя группа содержит медиальное преоптическое, супрахиазматическое, супраоптическое, паравентрикулярное и переднее гипоталамическое ядра;
средняя группа включает дорсо-медиальное, вентро- медиальное, аркуатное и латеральное гипоталамические ядра;
в состав задней группы входят супрамамиллярное, премамиллярное, мамиллярныеядра, задние гипоталамическое и перифорниатное ядра.
Важная физиологическая особенность гипоталамуса — высокая проницаемость его сосудов для различных веществ.
Гипоталамус тесно связан с деятельностью гипофиза. Средняя группа ядер образует медиальный гипоталамус и содержит нейроны- датчики, реагирующие на изменения состава и свойств внутренней среды организма. Латеральный гипоталамус формирует пути к верхним и нижним отделам ствола мозга.
Нейроны гипоталамуса получают импульсы с лимбической системы, ретикулярной формации, мозжечка, ядер таламуса, подкорковых ядер и коры; участвуют в оценке информации и формировании программы действий. Они имеют двусторонние связи с таламусом, а через него — с корой больших полушарий. Определенные нейроны гипоталамуса чувствительны к химическим воздействиям, гормонам, гуморальным факторам.
С передних ядер гипоталамуса осуществляются эфферентные влияния на исполнительные органы по парасимпатическому отделу, обеспечивающие общие парасимпатические приспособительные реакции (замедление сердечных сокращений, понижение тонуса сосудов и давления крови, увеличение секреции пищеварительных соков, усиление двигательной активности желудка и кишечника и др.). Через задние ядра осуществляются эфферентные влияния, поступающие к периферическим исполнительным органам по симпатическому отделу и обеспечивающие симпатические приспособительные реакции: учащение ритма сердечных сокращений, сужение сосудов и повышение давления крови, торможение моторной функции желудка и кишечника и др.
В передних и преоптических ядрах расположены высшие центры парасимпатического отдела, а в задних и латеральных ядрах — симпатического отдела нервной системы. Через эти центры обеспечивается интеграция соматических и вегетативных функций. В целом гипоталамус обеспечивает интеграцию деятельности эндокринной, вегетативной и соматической систем.
В латеральных ядрах гипоталамуса находится центр голода, ответственный за пищевое поведение. В медиальных ядрах расположен центр насыщения. Разрушение этих центров вызывает гибель животного. При раздражении центра насыщения прием корма прекращается, и возникают поведенческие реакции, характерные для состояния насыщения, а повреждение этого центра способствует повышенному потреблению корма и ожирению животных.
В средних ядрах находятся центры регуляции всех видов обмена веществ, энергорегуляции, теплорегуляции (теплообразования и теплоотдачи), половой функции, беременности, лактации, жажды.
Нейроны, расположенные в области супраоптического и пара- вентрикулярного ядер, участвуют в регуляции обмена воды. Раздражение их вызывает резкое увеличение потребления жидкости.
Гипоталамус является главной структурой, ответственной за температурный гомеостаз. В нем различают два центра: теплоотдачи и теплопродукции. Центр теплоотдачи локализован в передней и преоптической зонах гипоталамуса и включает паравентрикулярные, супраоптические и медиальные преоптические ядра. Раздражение этих структур вызывает увеличение теплоотдачи в результате расширения сосудов кожи и повышения температуры ее поверхности, увеличения потоотделения. Центр теплопродукции расположен в заднем гипоталамусе и состоит из различных ядер. Раздражение этого центра вызывает повышение температуры тела в результате усиления окислительных процессов, сужения сосудов кожи и появления мышечной дрожи.
Гипоталамус оказывает важное регулирующее влияние на половую функцию животных и человека.
Специфические ядра гипоталамуса (супраоптическое и паравентрикулярное) тесно взаимодействуют с гипофизом. Их нейроны секретируют нейрогормоны. В супраоптическом ядре образуется антидиуретический гормон (вазопрессин), в паравентрикулярном — окситоцин. Отсюда эти гормоны транспортируются по аксонам в гипофиз, где и накапливаются.
В нейронах гипоталамуса синтезируются либерины (рилизинг-гормоны) и статины, которые затем по нервным и сосудистым связям поступают в гипофиз. В гипоталамусе осуществляется интегрирование нервной и гуморальной регуляции функций многих органов. Гипоталамус и гипофиз образуют единую гипоталамо-гипофизарную систему с обратными связями. Уменьшение или увеличение количества гормонов в крови с помощью прямой и обратной афферентации изменяет активность нейросекреторных нейронов гипоталамуса, в результате чего изменяется уровень экскреции гипофизарных гормонов.
Подкорковые функции
Подкорковые функции в механизмах формирования поведенческих реакций человека и животных функции подкорковых образований проявляются всегда в тесном взаимодействии с корой больших полушарий. К подкорковым образованиям относят структуры, лежащие между корой и продолговатым мозгом: таламус (см. Головной мозг), гипоталамус (см.), базальные узлы (см.), комплекс образований, объединяемых в лимбическую систему мозга, а также ретикулярную формацию (см.) ствола мозга и таламуса. Последней принадлежит ведущая роль в формировании восходящих активирующих потоков возбуждения, генерализованно охватывающих кору больших полушарий. Любое афферентное возбуждение, возникшее при раздражении рецепторов на периферии, на уровне ствола мозга трансформируется в два потока возбуждений. Один поток по специфическим путям достигает специфической для данного раздражения проекционной области коры; другой — от специфического пути по коллатералям попадает в ретикулярную формацию и от нее в виде мощного восходящего возбуждения направляется к коре больших полушарий, активируя ее (рис.). Лишенная связей с ретикулярной формацией кора головного мозга приходит в недеятельное состояние, характерное для состояния сна.
Схема восходящего активирующего влияния ретикулярной формации (по Мегуну): 1 и 2 — специфический (лемнисковый) проводящий путь; 3 — коллатерали, отходящие от специфического пути к ретикулярной формации ствола мозга; 4 — восходящая активирующая система ретикулярной формации; 5 — генерализованное влияние ретикулярной формации на кору больших полушарий.
Ретикулярная формация имеет тесные функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом, лимбической системой, мозжечком, поэтому все наиболее общие функции организма (регуляция постоянства внутренней среды, дыхание, пищевая и болевая реакции) находятся в ее ведении. Ретикулярная формация является областью широкого взаимодействия потоков возбуждений различной природы, так как к ее нейронам конвергируют как афферентные возбуждения от периферических рецепторов (звуковых, световых, тактильных, температурных и др.), так и возбуждения, приходящие от других отделов головного мозга.
Афферентные потоки возбуждений от периферических рецепторов на пути к коре больших полушарий имеют многочисленные синаптические переключения в таламусе. От латеральной группы ядер таламуса (специфические ядра) возбуждения направляются по двум путям: к подкорковым ганглиям и к специфическим проекционным зонам коры мозга. Медиальная группа ядер таламуса (неспецифические ядра) служит местом переключения восходящих активирующих влияний, которые направляются от стволовой ретикулярной формации в кору мозга. Тесные функциональные взаимосвязи между специфическими и неспецифическими ядрами таламуса обеспечивают первичный анализ и синтез всех афферентных возбуждений, поступающих в головной мозг. У животных, находящихся на низких ступенях филогенетического развития, таламус и лимбические образования играют роль высшего центра интеграции поведения, обеспечивая все необходимые рефлекторные акты животного, направленные на сохранение его жизни. У высших животных и человека высшим центром интеграции является кора больших полушарий.
С функциональной точки зрения к подкорковым образованиям относят комплекс структур головного мозга, который играет ведущую роль в формировании основных врожденных рефлексов человека и животных: пищевых, половых и оборонительных. Этот комплекс получил название лимбической системы и включает в себя поясную извилину, гиппокамп, грушевидную извилину, обонятельный бугорок, миндалевидный комплекс и область перегородки. Центральное место среди образований лимбической системы отводится гиппокампу. Анатомически установлен гиппокампальный круг (гиппокамп → свод → мамиллярные тела → передние ядра таламуса → поясная извилина → cingulum → гиппокамп), который вместе с гипоталамусом играет ведущую роль в формировании эмоций. Регуляторные влияния лимбической системы широко распространяются на вегетативные функции (поддержание постоянства внутренней среды организма, регуляция кровяного давления, дыхания, тонуса сосудов, моторики желудочно-кишечного тракта, половых функций).
Кора больших полушарий оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. Существуют различные формы циклического взаимодействия между корой и подкоркой, выражающиеся в циркуляции возбуждений между ними. Наиболее выраженная замкнутая циклическая связь существует между таламусом и соматосенсорной областью коры мозга, составляющими в функциональном отношении единое целое. Корково-подкорковая циркуляция возбуждений определяется не только таламокортикальными связями, но и более обширной системой подкорковых образований. На этом базируется вся условно-рефлекторная деятельность организма. Специфика циклических взаимодействий коры и подкорковых образований в процессе формирования поведенческой реакции организма определяется его биологическими состояниями (голод, боль, страх, ориентировочно — исследовательская реакция).
Подкорковые функции. Кора головного мозга является местом высшего анализа и синтеза всех афферентных возбуждений, областью формирования всех сложных приспособительных актов живого организма. Однако полноценная аналитико-синтетическая деятельность коры больших полушарий возможна лишь при условии прихода к ней от подкорковых структур мощных генерализованных потоков возбуждений, богатых энергией и способных обеспечить системный характер корковых очагов возбуждений. С этой точки зрения и следует рассматривать функции подкорковых образований, являющихся, по выражению И. П. Павлова, «источником энергии для коры».
В анатомическом плане к подкорковым образованиям относят нейрональные структуры, расположенные между корой головного мозга (см.) и продолговатым мозгом (см.), а с функциональной точки зрения — подкорковые структуры, которые в тесном взаимодействии с корой больших полушарий формируют целостные реакции организма. Таковы таламус (см.), гипоталамус (см.), базальные узлы (см.), так называемая лимбическая система мозга. С функциональной точки зрения к подкорковым образованиям относят и ретикулярную формацию (см.) ствола мозга и таламуса, которой принадлежит ведущая роль в формировании восходящих активирующих потоков к коре больших полушарий. Восходящие активирующие влияния ретикулярной формации открыли Моруцци и Мегун (G. Moruzzi, Н. W. Magoun). Раздражая электрическим током ретикулярную формацию, эти авторы наблюдали переход медленной электрической активности коры головного мозга в высокочастотную, низкоамплитудную. Такие же изменения электрической активности коры мозга («реакция пробуждения», «реакция десинхронизации») наблюдались при переходе от сонного состояния животного к бодрствующему. На основании этого возникло предположение о пробуждающем влиянии ретикулярной формации (рис. 1).
Рис. 1. «Реакция десинхронизации» корковой биоэлектрической активности при раздражении у кошки седалищного нерва (отмечено стрелками): СМ — сенсомоторная область коры мозга; ТЗ — теменно-затылочная область коры мозга (л — левая, п — правая).
В настоящее время известно, что реакция десинхронизации корковой электрической активности (активация коры мозга) может возникать при любом афферентном воздействии. Это связано с тем, что на уровне ствола мозга афферентное возбуждение, возникающее при раздражении любых рецепторов, трансформируется в два потока возбуждения. Один поток направляется по классическому лемнисковому пути и достигает специфической для данного раздражения корковой проекционной области; другой — попадает от лемнисковой системы по коллатералям в ретикулярную формацию и от нее в виде мощных восходящих потоков направляется к коре мозга, генерализованно активируя ее (рис. 2).
Рис. 2. Схема восходящего активирующего влияния ретикулярной формации (по Мегуну): 1—3 — специфический (лемнисковый) проводящий путь; 4 — коллатерали, отходящие от специфического пути к ретикулярной формации ствола мозга; 5 — восходящая активирующая система ретикулярной формации; в — генерализованное влияние ретикулярной формации на кору больших полушарий.
Это генерализованное восходящее активирующее влияние ретикулярной формации — непременное условие поддержания бодрствующего состояния мозга. Лишенная источника возбуждения, которым служит ретикулярная формация, кора головного мозга приходит в недеятельное состояние, сопровождаемое медленной высокоамплитудной электрической активностью, характерной для состояния сна. Такую картину можно наблюдать при децеребрации, т. е. у животного с перерезанным стволом мозга (см. ниже). В этих условиях ни какие-либо афферентные раздражения, ни прямое раздражение ретикулярной формации не вызывает диффузной, генерализованной реакции десинхронизации. Таким образом, доказано наличие в головном мозге по крайней мере двух основных каналов поступления афферентных воздействий на кору больших полушарий: по классическому лемнисковому пути и по коллатералям через ретикулярную формацию ствола мозга.
Так как при любом афферентном раздражении генерализованная активация коры мозга, оцениваемая по электроэнцефалографическому показателю (см. Электроэнцефалография), всегда сопровождается реакцией десинхронизации, многие исследователи пришли к выводу, что любые восходящие активирующие влияния ретикулярной формации на кору головного мозга являются неспецифическими. Главными аргументами в пользу такого вывода явились следующие: а) отсутствие сенсорной модальности, т. е. однотипность изменений биоэлектрической активности при воздействии различных сенсорных раздражителей; б) постоянный характер активации и генерализованное распространение возбуждения по коре, оцениваемое опять-таки по электроэнцефалографическому показателю (реакция десинхронизации). На этом основании все виды генерализованной десинхронизации корковой электрической активности признавались также едиными, не различающимися по каким-либо физиологическим качествам. Однако при формировании целостных приспособительных реакций организма восходящие активирующие влияния ретикулярной формации на кору головного мозга носят специфический характер, соответствующий данной биологической деятельности животного — пищевой, половой, оборонительной (П. К. Анохин). Это означает, что в формировании различных биологической реакций организма участвуют различные области ретикулярной формации, осуществляющие активацию коры больших полушарий (А. И. Шумилина, В. Г. Агафонов, В. Гавличек).
Наряду с восходящими влияниями на кору больших полушарий ретикулярная формация может оказывать и нисходящие влияния на рефлекторную деятельность спинного мозга (см.). В ретикулярной формации различают области, которые оказывают тормозящие и облегчающие влияния на моторную активность спинного мозга. По своему характеру эти влияния диффузны и оказывают воздействие на все группы мышц. Они передаются по нисходящим спинальным путям, которые различны для тормозящих и облегчающих влияний. О механизме ретикулоспинальных влияний существует две точки зрения: 1) ретикулярная формация оказывает тормозящие и облегчающие влияния непосредственно на мотоневроны спинного мозга; 2) эти влияния на мотоневроны передаются через клетки Реншо. Особенно четко нисходящие влияния ретикулярной формации выражены у децеребрированного животного. Децеребрация осуществляется путем перерезки головного мозга по передней границе четверохолмия. При этом развивается так называемая децеребрационная ригидность с резким увеличением тонуса всех мышц-разгибателей. Считают, что этот феномен развивается в результате перерыва путей, идущих от вышележащих образований мозга к тормозящему отделу ретикулярной формации, что обусловливает уменьшение тонуса этого отдела. В результате облегчающие влияния ретикулярной формации начинают преобладать, что и приводит к увеличению тонуса мышц.
Важной особенностью ретикулярной формации является ее высокая чувствительность к различным химическими веществам, циркулирующим в крови (СO2, адреналин и др.). Это обеспечивает включение ретикулярной формации в регулирование некоторых вегетативных функций. Ретикулярная формация является также местом избирательного действия многих фармакологических и медикаментозных препаратов, что используют при лечении некоторых заболеваний ЦНС. Высокая чувствительность ретикулярной формации к барбитуратам и ряду нейроплегических средств позволила по-новому представить механизм наркотического сна. Действуя тормозящим образом на нейроны ретикулярной формации, наркотик тем самым лишает кору головного мозга источника активирующих влияний и обусловливает развитие состояния сна. Гипотермическое действие аминазина и подобных ему препаратов объясняют влиянием этих веществ на ретикулярную формацию.
Ретикулярная формация имеет тесные функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом и другими отделами головного мозга, поэтому все наиболее общие функции организма (терморегуляция, пищевая и болевая реакции, регуляция постоянства внутренней среды организма) находятся в той или иной функциональной зависимости от нее. Ряд исследований, сопровождавшийся регистрацией при помощи микроэлектродной техники электрической активности отдельных нейронов ретикулярной формации, показал, что эта область является местом взаимодействия афферентных потоков различной природы. К одному и тому же нейрону ретикулярной формации могут конвергировать возбуждения, возникающие не только при раздражении различных периферических рецепторов (звуковых, световых, тактильных, температурных и др.), но и идущие от коры больших полушарий, мозжечка и других подкорковых структур. На основе этого механизма конвергенции в ретикулярной формации происходит перераспределение афферентных возбуждений, после чего они в виде восходящих активирующих потоков направляются к нейронам коры головного мозга.
Прежде чем достигнуть коры, эти потоки возбуждения имеют многочисленные синаптические переключения в таламусе, который служит как бы промежуточным, связующим звеном между низшими образованиями ствола мозга и корой больших полушарий. Импульсы от периферических концов всех внешних и внутренних анализаторов (см.) переключаются в латеральной группе ядер таламуса (специфические ядра) и отсюда направляются по двум путям: к подкорковым ганглиям и к специфическим проекционным зонам коры мозга. Медиальная группа ядер таламуса (неспецифические ядра) служит местом переключения восходящих активирующих влияний, которые направляются от стволовой ретикулярной формации в кору мозга.
Специфические и неспецифические ядра таламуса находятся в тесной функциональной взаимосвязи, что обеспечивает первичный анализ и синтез всех афферентных возбуждений, поступающих в головной мозг. В таламусе имеется четкая локализация представительства различных афферентных нервов, идущих от различных рецепторов. Эти афферентные нервы заканчиваются в определенных специфических ядрах таламуса, а от каждого ядра волокна направляются в кору головного мозга к специфическим проекционным зонам представительства той или иной афферентной функции (зрительной, слуховой, тактильной и т. д.). Особенно тесно таламус связан с соматосенсорной областью коры больших полушарий. Эта взаимосвязь осуществляется благодаря наличию замкнутых циклических связей, направленных как от коры к таламусу, так и от таламуса к коре. Поэтому соматосенсорную область коры и таламус в функциональном отношении можно рассматривать как единое целое.
У животных, находящихся на более низких ступенях филогенетического развития, таламус играет роль высшего центра интеграции поведения, обеспечивая все необходимые рефлекторные акты животного, направленные на сохранение его жизни. У животных, стоящих на высших ступенях филогенетической лестницы, и у человека высшим центром интеграции становится кора больших полушарий. Функции же таламуса заключаются в регуляции и осуществлении ряда сложных рефлекторных актов, являющихся как бы базой, на основе которой создается адекватное целенаправленное поведение животного и человека. Эти ограниченные функции таламуса четко проявляются у так называемого таламического животного, т. е. у животного с удаленными корой больших полушарий и подкорковыми узлами. Такое животное может самостоятельно передвигаться, сохраняет основные позно-тонические рефлексы, обеспечивающие нормальное положение тела и головы в пространстве, сохраняет регуляцию температуры тела и всех вегетативных функций. Но оно не может адекватно реагировать на различные раздражители внешней среды вследствие резкого нарушения условно-рефлекторной деятельности. Таким образом, таламус в функциональной взаимосвязи с ретикулярной формацией, оказывая локальные и генерализованное воздействия на кору больших полушарий, организует и регулирует соматическую функцию головного мозга как целого.
Среди структур головного мозга, относящихся к подкорковым с функциональной точки зрения, выделяют комплекс образований, который играет ведущую роль в формировании основных врожденных активностей животного: пищевой, половой и оборонительный. Этот комплекс получил название лимбической системы мозга и включает в себя гиппокамп, грушевидную извилину, обонятельный бугорок, миндалевидный комплекс и область перегородки (рис. 3). Все эти образования объединяются на функциональной основе, так как они принимают участие в обеспечении поддержания постоянства внутренней среды, регуляции вегетативных функций, в формировании эмоций (см.) и мотиваций (см.). Многие исследователи относят к лимбической системе и гипоталамус. Лимбическая система принимает непосредственное участие в формировании эмоционально окрашенных, примитивных врожденных форм поведения. Особенно это относится к формированию половой функции. При поражении (опухоль, травма и др.) некоторых структур лимбической системы (височная область, поясная извилина) у человека нередко наблюдаются сексуальные расстройства.
Рис. 3. Схематическое изображение основных связей лимбической системы (по Мак-Лейну): N — nucleus interpeduncularis; MS и LS — медиальная и латеральная обонятельные полоски; S — перегородка; MF — медиальный пучок переднего мозга; Т — обонятельный бугорок; AT — переднее ядро таламуса; М — мамиллярное тело; SM — stria medialis (стрелками обозначено распространение возбуждения по лимбической системе).
Центральное место среди образований лимбической системы отводится гиппокампу. Анатомически установлен гиппокампальный круг (гиппокамп → свод → мамиллярные тела → передние ядра таламуса → поясная извилина → cingulum → гиппокамп), который вместе с, гипоталамусом (си.) играет ведущую роль в формировании эмоций. Непрерывная циркуляция возбуждения по гиппокампальному кругу определяет главным образом тоническую активацию коры головного мозга, а также интенсивность эмоций.
Часто у больных с тяжелыми формами психоза и другими психическими заболеваниями после смерти находили патологические изменения в структурах гиппокампа. Предполагают, что циркуляция возбуждения по гиппокампальному кольцу служит одним из механизмов памяти. Отличительная особенность лимбической системы — тесная функциональная взаимосвязь между ее структурами. Благодаря этому возбуждение, возникшее в какой-либо структуре лимбической системы, тут же охватывает остальные образования и долгое время не выходит за пределы всей системы. Подобное длительное, «застойное» возбуждение лимбических структур, вероятно, также лежит в основе формирования эмоциональных и мотивационных состояний организма. Некоторые образования лимбической системы (миндалевидный комплекс) оказывают генерализованное восходящее активирующее влияние на кору головного мозга.
Учитывая регуляторные влияния лимбической системы на вегетативные функции (кровяное давление, дыхание, тонус сосудов, моторику желудочно-кишечного тракта), можно понять те вегетативные реакции, которые сопровождают любой условнорефлекторный акт организма. Этот акт как целостная реакция осуществляется всегда при непосредственном участии коры больших полушарий, которая является высшей инстанцией анализа и синтеза афферентных возбуждений. У животных после удаления коры головного мозга (декортицированных) резко нарушается условно-рефлекторная деятельность, причем, чем выше стоит животное в эволюционном отношении, тем ярче выражены эти нарушения. Поведенческие реакции животного, подвергшегося декортикации, сильно расстраиваются; большую часть времени такие животные спят, просыпаясь только при сильных раздражениях и для совершения простых рефлекторных актов (мочеиспускание, дефекация). У таких животных можно выработать условнорефлекторные реакции, однако слишком примитивные и недостаточные для осуществления адекватной приспособительной деятельности организма.
Вопрос о том, на каком уровне головного мозга (в коре или подкорке) происходит замыкание условного рефлекса, в настоящее время не рассматривается как принципиальный. Мозг участвует в формировании приспособительного поведения животного, в основе которого лежит принцип условного рефлекса, как единая целостная система. Любые раздражители — как условные, так и безусловные — конвергируют к одному и тому же нейрону различных подкорковых образований, а также к одному нейрону различных областей коры больших полушарий. Изучение механизмов взаимодействия коры и подкорковых образований в процессе формирования поведенческой реакции организма — одна из основных задач современной физиологии головного мозга. Кора больших полушарий, являясь высшей инстанцией синтеза афферентных возбуждений, организует внутренние нервные связи для совершения ответного рефлекторного акта. Ретикулярная формация и другие подкорковые структуры, оказывая множественные восходящие влияния на кору головного мозга, создают лишь необходимые условия для организации более совершенных корковых временных связей, а в результате этого — и для формирования адекватной поведенческой реакции организма. Кора больших полушарий в свою очередь оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. В этом тесном функциональном взаимодействии между корой и нижележащими образованиями головного мозга заключена основа интегративной деятельности мозга как единого целого. С этой точки зрения, разделение функций мозга на чисто корковые и чисто подкорковые в какой-то степени искусственно и необходимо лишь для понимания роли различных образований мозга в формировании целостной приспособительной реакции организма.
Промежуточный мозг Анатомия промежуточного мозга
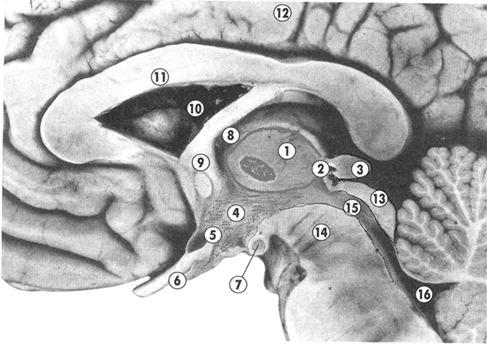
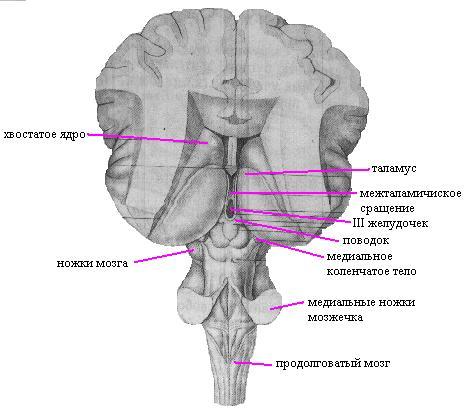
Промежуточный мозг располагается под мозолистым телом и сводом, срастаясь по бокам с полушариями большого мозга. К нему относятся: таламус (зрительные бугры), эпиталамус (надбугорная область), метаталамус (забугорная область) и гипоталамус (подбугорная область). Полостью промежуточного мозга является III желудочек.
Таламус представляет собой парные скопления серого вещества, покрытые слоем белого вещества, имеющие яйцевидную форму. Передний отдел его примыкает к межжелудочковому отверстию, задний, расширенный, — к четверохолмию. Латеральная поверхность таламуса срастается с полушариями и граничит с хвостатым ядром и внутренней капсулой. Медиальные поверхности обр азуют стенки III желудочка. Нижняя продолжается в гипоталамус. В таламусе различают три основные группы ядер: передние, латеральные и медиальные. В латеральных ядрах происходит переключение всех чувствительных путей, направляющихся к коре больших полушарий. В эпиталамусе лежит верхний придаток мозга — эпифиз, или шишковидное тело, подвешенное на двух поводках в углублении между верхними холмиками пластинки крыши. Метаталамус представлен медиальными и латеральными коленчатыми телами, соединенными пучками волокон (ручки холмиков) с верхними (латеральные) и нижними (медиальные) холмиками пластинки крыши. В них лежат ядра, являющиеся рефлекторными центрами зрения и слуха.
Гипоталамус располагается вентральнее зрительного бугра и включает в себя собственно подбугорную область и ряд образований, расположенных на основании мозга. Сюда относятся; конечная пластинка, зрительный перекрест, серый бугор, воронка с отходящим от нее нижним придатком мозга — гипофизом и сосцевидные тела. В гипоталамической области расположены ядра (надзрительное, околожелудочковое и др.), содержащие крупные нервные клетки, способные выделять секрет (нейросекрет), поступающий по их аксонам в заднюю долю гипофиза, а затем в кровь. В заднем отделе гипоталамуса лежат ядра, образованные мелкими нервными клетками, которые связаны с передней долей гипофиза особой системой кровеносных сосудов.
Третий желудочек расположен по средней линии и представляет собой узкую вертикальную щель. Боковые стенки его образованы зрительными буграми и подбугорной областью, передняя — столбами свода и передней спайкой, нижняя — образованиями гипоталамуса и задняя — ножками мозга и надбугорной областью. Верхняя стенка — крыша III желудочка, — самая тонкая и состоит из мягкой (сосудистой) оболочки мозга, выстланной со стороны полости желудочка эпителиальной пластинкой (эпендимой). Отсюда в полость желудочка вдавливается большое количество кровеносных сосудов: и образуется сосудистое сплетение. Спереди III желудочек сообщается с боковыми желудочками (I и II) межжелудочковыми отверстиями, а сзади переходит в мозговой водопровод.
Рис 5. Ствол головного мозга, вид сверху и сзади .
Физиология промежуточного мозга.
Главными образованиями промежуточного мозга являются таламус (зрительный бугор) и гипоталамус (подбугорная область).
Таламус — чувствительное ядро подкорки. Его называют «коллектором чувствительности», так как к нему сходятся афферентные (чувствительные) пути от всех рецепторов, исключая обонятельные. Здесь находится третий нейрон афферентных путей, отростки которого заканчиваются в чувствительных зонах коры.
Главной функцией таламуса является интеграция (объединение) всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. Здесь происходит сопоставление информации, получаемой по различным каналам связи, и оценка ее биологического значения. В зрительном бугре насчитывается 40 пар ядер, которые подразделяются на специфические (на нейронах этих ядер заканчиваются восходящие афферентные пути), неспецифические (ядра ретикулярной формации) и ассоциативные. Через ассоциативные ядра таламус связан со всеми двигательными ядрами подкорки — полосатым телом, бледным шаром, гипоталамусом и с ядрами среднего и продолговатого мозга.
Изучение функций зрительного бугра проводится путем перерезок, раздражения и разрушения.
Кошка, у которой разрез сделан выше промежуточного мозга, резко отличается от кошки, у которой высшим отделом центральной нервной системы является средний мозг. Она не только поднимается и ходит, т. е. выполняет сложно координированные движения, но еще проявляет все признаки эмоциональных реакций. Легкое прикосновение вызывает злобную реакцию. Кошка бьет хвостом, скалит зубы, рычит, кусается, выпускает когти. У человека зрительный бугор играет существенную роль в эмоциональном поведении, характеризующемся своеобразной мимикой, жестами и сдвигами функций внутренних органов. При эмоциональных реакциях повышается давление, учащаются пульс, дыхание, расширяются зрачки. Мимическая реакция человека является врожденной. Если пощекотать нос плода 5 — 6 мес., то можно видеть типичную гримасу неудовольствия (П. К. Анохин). При раздражении зрительного бугра у животных возникают двигательные и болевые реакции — визг, ворчание. Эффект можно объяснить тем, что импульсы от зрительных бугров легко переходят на связанные с ними двигательные ядра подкорки.
В клинике симптомами поражения зрительных бугров являются сильная головная боль, расстройства сна, нарушения чувствительности как в сторону повышения, так и понижения, нарушения движений, их точности, соразмерности, возникновение насильственных непроизвольных движений.
Гипоталамус является высшим подкорковым центром вегетативной нервной системы. В этой области расположены центры, регулирующие все вегетативные функции, обеспечивающие постоянство внутренней среды организма, а также регулирующие жировой, белковый, углеводный и водно-солевой обмен.
В деятельности вегетативной нервной системы гипоталамус играет такую же важную роль, какую играют красные ядра среднего мозга в регуляции скелетно-моторных функций соматической нервной системы.
Самые ранние исследования функций гипоталамуса принадлежат — Клоду Бернару. Он обнаружил, что укол в промежуточный мозг кролика вызывает повышение температуры тела почти на 3°С. Этот классический опыт, открывший локализацию центра терморегуляции в гипоталамусе, получил название теплового укола. После разрушения гипоталамуса животное становится пойкилотермным, т. е. теряет способность удерживать постоянство температуры тела. В холодной комнате температура тела понижается, а в жаркой повышается.
Позднее было установлено, что почти все органы, иннервируемые вегетативной нервной системой, могут быть активированы раздражением подбугорной области. Иными словами, все эффекты, которые можно получить при раздражении симпатических и парасимпатических нервов, получаются при раздражении гипоталамуса.
В настоящее время для раздражения различных структур мозга широко применяется метод вживления электродов. С помощью особой, так называемой стереотаксической техники, через трепанационное отверстие в черепе вводят электроды в любой заданный участок мозга. Электроды изолированы на всем протяжении, свободен только их кончик. Включая электроды в цепь, можно узко локально раздражать те или иные зоны.
При раздражении передних отделов гипоталамуса возникают парасимпатические эффекты — усиление движений кишечника, отделение пищеварительных соков, замедление сокращений сердца и др. при раздражении задних отделов наблюдаются симпатические эффекты — учащение сердцебиения, сужение сосудов, повышение температуры тела и др. Следовательно, в передних отделах подбугорной области располагаются парасимпатические центры, а в задних — симпатические.
Так как раздражение при помощи вживленных электродов производится на целом животном, без наркоза, то представляется возможность судить о поведении животного. В опытах Андерсена на козе с вживленными электродами был найден центр, раздражение которого вызывает неутолимую жажду, — центр жажды. При его раздражении коза могла выпивать до 10 л воды. Раздражением других участков можно было заставить сытое животное есть (центр голода).
Широкую известность получили опыты испанского ученого Дельгадо на быке с электродом, вживленным в центр страха: Когда на арене разъяренный бык бросался на тореадора, включали раздражение, и бык отступал с ясно выраженными признаками страха.
Американский исследователь Д. Олдз предложил модифицировать метод — предоставить возможность самому животному замыкать, что неприятных раздражений животное будет избегать и, наоборот, стремиться повторять приятные.
Опыты показали, что имеются структуры, раздражение которых вызывает безудержное стремление к повторению. Крысы доводили себя до истощения, нажимая на рычаг до 14000 раз! Кроме того, обнаружены структуры, раздражение которых, по-видимому, вызывает крайне неприятное ощущение, так как крыса второй раз избегает нажать на рычаг и убегает от него. Первый центр, очевидно, является центром удовольствия, второй — центром неудовольствия.
Чрезвычайно важным для понимания функций гипоталамуса явилось открытие в этом отделе мозга рецепторов, улавливающих изменения температуры крови (терморецепторы), осмотического давления (осморецепторы) и состава крови (глюкорецепторы).
С рецепторов, обращенных в кровь, возникают рефлексы, направленные на поддержание постоянства внутренней среды организма — гомеостаза. «Голодная кровь», раздражая глюкорецепторы, возбуждает пищевой центр: возникают пищевые реакции, направленные на поиск и поедание пищи.
Одним из частых проявлений заболевания гипоталамуса в клинике является нарушение водно-солевого обмена, проявляющееся в выделении большого количества мочи с низкой плотностью. Заболевание носит название несахарного мочеизнурения.
Подбугорная область тесно связана с деятельностью гипофиза. В крупных нейронах надзрительного и околожелудочкового ядер гипоталамуса образуются гормоны — вазопрессин и окситоцин. По аксонам гормоны стекают к гипофизу, где накапливаются, а затем поступают в кровь.
Иное взаимоотношение между гипоталамусом и передней долей гипофиза. Сосуды, окружающие ядра гипоталамуса, объединяются в систему вен, которые спускаются к передней доле гипофиза и здесь распадаются на капилляры. С кровью к гипофизу поступают вещества — релизинг-факторы, или освобождающие факторы, стимулирующие образование гормонов в передней его доле.
Ретикулярная формация. В стволе мозга — продолговатом, среднем и промежуточном мозге, между его специфическими ядрами находятся скопления нейронов с многочисленными сильно ветвящимися отростками, образующими густую сеть. Эта система нейронов получила название сетчатого образования, или ретикулярной формации. Специальные исследования показали, что все так называемые специфические пути, проводящие определенные виды чувствительности от рецепторов к чувствительным зонам коры головного Мозга, дают в стволе мозга ответвления, заканчивающиеся на клетках ретикулярной формации. Потоки импульсов с периферии от экстеро-, интеро- и проприорецепторов. поддерживают постоянное тоническое возбуждение структур ретикулярной формации.
От нейронов ретикулярной формации начинаются неспецифические пути. Они поднимаются вверх к коре головного мозга и подкорковым ядрам и спускаются вниз к нейронам спинного мозга.
В чем же состоит функциональное значение этой своеобразной системы, не имеющей своей территории, располагающейся между специфическими соматическими и вегетативными ядрами ствола мозга?
Методом раздражения отдельных структур ретикулярной формации удалось раскрыть ее функцию как регулятора функционального состояния спинного и головного мозга, а также важнейшего регулятора мышечного тонуса. Роль ретикулярной формации в деятельности центральной нервной системы сравнивают с ролью регулятора в телевизоре. Не давая изображения, он может менять громкость звука и освещенность.
Раздражение ретикулярной формации, не вызывая двигательного эффекта, изменяет имеющуюся деятельность, тормозя ее или усиливая. Если у кошки короткими, ритмическими раздражениями чувствительного нерва вызывать защитный рефлекс — сгибание задней лапки, а затем на этом фоне присоединить раздражение ретикулярной формации, то в зависимости от зоны раздражения эффект будет различен: спинальные рефлексы либо резко усилятся, либо ослабятся и исчезнут, т. е. затормозятся. Торможение возникает при раздражении задних отделов ствола мозга, а усиление рефлексов — при раздражении передних отделов. Соответствующие зоны ретикулярной формации получили название тормозящей и активирующей зон.
На кору головного мозга ретикулярная формация оказывает активирующее воздействие, поддерживая состояние бодрствования и концентрируя внимание. Если у спящей кошки с вживленными в промежуточный мозг электродами включать раздражение ретикулярной формации, то кошка просыпается, открывает глаза. На электроэнцефалограмме видно, что исчезают медленные волны, характерные для сна, и появляются быстрые волны, свойственные состоянию бодрствования. Ретикулярная формация оказывает на кору головного мозга восходящее, генерализованное (охватывающее всю кору) активирующее влияние. По выражению И.П. Павлова, «подкорка заряжает кору». В свою очередь кора больших полушарий регулирует активность сетчатого образования.
Четверохолмие: подкорковые центры зрительного и слухового анализаторов. Рефлексы четверохолмия
Крыша среднего мозга (пластинка четверохолмия). Важным отделом среднего мозга является четверохолмие, состоящее из двух передних и двух задних бугров, а также пластинки крыши, на которой эти холмики расположены. Внутри холмиков имеется ядра.
· Ядра передних холмиков представляют собой первичные зрительные центры, обеспечивающие формирование зрачковых и зрительных ориентировочных рефлексов (на внезапные световые раздражители). Они получают сигналы непосредственно от сетчатки глаза и являются первичными подкорковыми центрами зрения.
· а задние первичные слуховые центры, получают информацию от ядер слуховых нервов. К ориентировочным слуховым рефлексам относятся – рефлекс настораживания (поворот ушей и головы и тела к источнику звука). Здесь расположен первичный подкорковый центр слуха.
Ориентировочные рефлексы играют большую роль в жизни животных и человека. Они лежат в основе реакций, благодаря которым субъект может мгновенно включаться в действие, требующее быстроты, внезапности.
От каждого холмика в сторону отходит валик белого вещества, который называется ручки холмика.
· Ручки верхнего холмика следуют к латеральному коленчатому телу
· Ручки нижнего холмика – к медиальному коленчатому телу (коленчатые тела относятся к промежуточному мозгу)
Коленчатые тела относятся к таламическому отделу промежуточного мозга. Здесь расположены вторичные подкорковые центры зрения и слуха.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения:Увлечёшься девушкой-вырастут хвосты, займёшься учебой-вырастут рога 8913 — | 6984 — или читать все.
Отключите adBlock! и обновите страницу (F5) очень нужно
Тема 12. Промежуточный мозг.
Промежуточный мозг в процессе эмбриогенеза развивается из переднего мозгового пузыря. Он образует стенки третьего мозгового желудочка. В состав промежуточного мозга входят: зрительные бугры (таламус), гипоталамус, эпиталамус и метаталамус.
Таламус (зрительные бугры) представляет собой скопление серого вещества, имеющего яйцевидную форму. Его длина составляет около 40 мм, ширина 16 мм, высота 20 мм. Медиальная и дорсальная поверхности свободны, вентральная и латеральная поверхности сращены со структурами конечного мозга. Медиальная поверхность таламуса обращена в полость III желудочка.
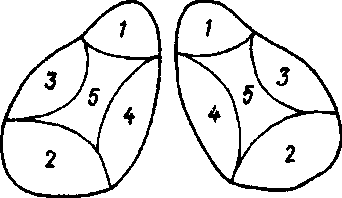
Таламус является крупным подкорковым образованием, через которое в кору больших полушарий проходят разнообразные афферентные пути. Нервные клетки его группируются в большое количество ядер (около120). Топографически последние разделяют на переднюю, заднюю, срединную, медиальную и латеральную группы:
Передние ядра таламуса являются подкорковым центром обоняния.
Задние ядра – подкорковым центром зрения.
Срединные ядра – подкорковые центры вестибулярных и слуховых функций.
Медиальные ядра — интегративным центром промежуточного мозга, в который поступает информация от остальных ядер таламуса, а также центром экстрапирамидной системы.
Латеральные ядра – подкорковый центр общей чувствительности.
Рис. 2. Группы ядер таламуса
1 — передняя группа (обонятельные); 2 — задняя группа (зрительные); 3 — латеральная группа (общая чувствительность); 4 — медиальная группа (экстрапирамидная система, интегративный центр таламуса); 5 – срединнаягруппа (вестибулярных и слуховых функций).
По функции таламические ядра можно дифференцировать на специфические, неспецифические и ассоциативные.
Специфические ядра делятся на чувствительные и двигательные. От чувствительных ядер информация о характере сенсорных стимулов поступает в строго определенные участки 3-4 слоев коры. Чувствительные ядра имеют топическую организацию, строго организованную локальную систему входов и выходов. Функциональной единицей специфических чувствительных таламических ядер являются «релейные» нейроны, которые имеют мало дендритов, длинный аксон и выполняют переключательную функцию. Здесь происходит переключение путей, идущих в кору от кожной, мышечной и других видов чувствительности. Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности. К двигательным ядрам таламуса относится вентральное ядро, которое имеет вход от мозжечка и базальных ганглиев, и одновременно дает проекции в моторную зону коры больших полушарий. Это ядро включено в систему регуляции движений.
Неспецифические ядра таламуса имеют обширные входы от разных отделов мозга, связаны с обширными участками коры и принимают участие в активизации ее деятельности, их относят к ретикулярной формации. Неспецифический ядра таламуса принимают участие в организации процессов внимания.
Ассоциативные ядра образованы мультиполярными и биполярными нейронами. Ядра не имеют прямых контактов с афферентными системами. Получают импульсы от релейных ядер таламуса. От них импульсы идут в кору головного мозга в ассоциативные зоны (в третичные проекционные зоны), за счет этих импульсов возникают примитивные ощущения; также они обеспечивают взаимосвязь между сенсорными системами в коре головного мозга.
Функционально таламус – структура, в которой происходит обработка и интеграция практически всех сенсорных сигналов, идущих в кору головного мозга. Возможность получить информацию о состоянии множества систем организма позволяет ему участвовать в регуляции и определять функциональное состояние организма в целом. В связи с этим таламус фактически является подкорковым чувствительным центром. Отростки нейронов таламуса направляются отчасти к ядрам полосатого тела конечного мозга (в связи с этим таламус рассматривается как чувствительный центр экстропирамидной системы), отчасти к коре большого мозга, образуя таламокортикальные пути.
Гипоталамус (подталамическая область), hypothalamus, — сравнительно небольшое, но исключительно важное образование головного мозга, образующее нижние отделы промежуточного мозга. Гипоталамус спереди граничит со зрительным перекрестом; задняя граница — сосцевидные тела, латерально ограничен зрительными трактами. Верхней границей является гипоталамическая борозда. Нижняя граница, или дно III желудочка, представлена серым бугром, лежащим впереди сосцевидных тел. Нижняя стенку III желудочка кпереди продолжается в концевую пластинку конечного мозга.
Учитывая, что гипоталамус включает большое количество отдельных образований, целесообразно сгруппировать их по топографическому принципу следующим образом.
1. Передняя гипоталамическая область, или зрительная часть:
— зрительный перекрест, chiasma optictnn;
— зрительный тракт, tractus opticus.
П. Промежуточная гипоталамическая область:
— серый бугор, tuber cinereum;
III. Задняя гипоталамическая область, или сосочковая часть,
— заднее гипоталамическое ядро;
— сосочковые тела, corpora mamillaria.
Рассмотрим внешнюю форму этих образований:
Зрительная часть гипоталамуса включает перекрест зрительных нервов, правый и левый зрительные тракты. Зрительный перекрест имеет вид поперечно лежащего валика, образованного волокнами зрительных нервов, которые здесь частично переходят на противоположную сторону. От задних углов зрительного перекреста отходят зрительные тракты, имеющие вид белых тяжей, сращенных с веществом мозга. Они идут латерально и назад, огибают ножки мозга и заканчиваются двумя корешками. Более крупный латеральный корешок заканчивается в латеральных коленчатых телах, а медиальный – направляется к верхнему холму четверохолмия.
Кзади от зрительного перекреста находится серый бугор.Серый 6угop, tuber cinereum,— тонкостенная часть дна III желудочка, расположенная между сосцевидными телами и перекрестом зрительного нерва. Стенки серого бугра образованы тонкой пластинкой серого вещества. Кпереди серый бугор переходит в истонченную конечную пластинку (lamina terminalis). Она натянута между перекрестом зрительных нервов и передней мозговой спайкой. В ядрах серого бугра расположены высшие центры регуляции вегетативного отдела нервной системы. Серый бугор вытягивается в воронку (infundibulum), на которой висит гипофиз.
Гипофиз(hypophysis), или нижний придаток мозга имеет яйцевидную форму, соединен ножкой с серым бугром промежуточного мозга. Он имеет форму фасоли массой 0,4-0,6 г, размером 10х12х6 мм. У женщин, особенно беременных, гипофиз несколько больше: масса его иногда достигает 1,0-1,2 г, обычно 0,7 г. Гипофиз состоит из трех долей — задней, промежуточной и передней, окруженных общей соединительнотканной оболочкой. Задняя доля, меньшего размера, посредством ножки связана с воронкой. Между передней и задними долями имеется небольших размеров промежуточная доля, отделенная щелью от передней доли. Передняя доля непосредственной связи с головным мозгом не имеет, так как она имеет другое происхождение. Она представляет собой выпячивание эпителия первичной ротовой ямки. Задняя доля образуется путем выпячивания вентральной стенки промежуточного мозга.
Сосочковые тела, corpora mamillaria, — сферической формы, диаметром 5-6 мм, белого цвета. Они располагаются между серым бугром и задним продырявленным пространством. Белое вещество только снаружи, внутри находится серое вещество. Вместе с передними ядрами таламуса их рассматривают как подкорковые центры обоняния.
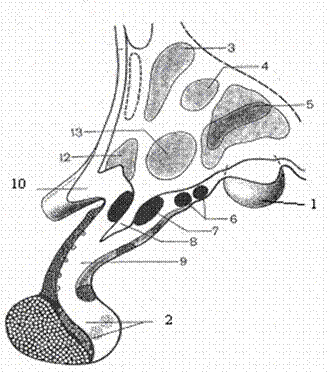
Пути и центры гипоталамуса. Ядра подталамической области, весьма многочисленные (около 40), располагаются главным образом в собственно подталамической области. По расположению их разделяют на три группы: переднюю, промежуточную и заднюю.
1. Передняя группа ядер включает супраоптическое, предоптическое и паравентрикулярные ядра. Эти ядра являются нейросекреторными. Надзрительное ядро (nucl supraopticus) — парное, располагается латеральнее медиальной плоскости над зрительным трактом от начала перекреста зрительного тракта и распространяется до середины серого бугра; клетки этого ядра вырабатывают антидиуретические гормоны (вазопрессин). Вазопрессин выделяется в ответ на раздражение, идущее с осморецепторов. Надзрительное ядро совместно с околожелудочковым ядром вырабатывает также нейрофизины — белки-носители. Околожелудочковое ядро (nucl. paraventricularis) — парное, в виде пластинки, располагается несколько выше III желудочка. Нижняя его часть начинается на уровне перекреста зрительных нервов, затем идет вверх и назад. Паравентрикулярные (околожелудочковые) ядра вырабатывают гормон окситоцин в ответ на раздражение механорецепторов матки и молочных желез. Отростки нейросекреторных клеток надзрительного и околожелудочкового ядер образуют гипоталамо-гипофизарный пучок, по аксонам которого их нейросекрет стекает в заднюю долю гипофиза в тельца скопления нейросекрета. В последних образуются гормоны задней доли гипофиза — антидиуретический гормон (АДГ) и окситоцин.
2. Промежуточная группа представлена ядрами собственно подталамической области, ядрами серого бугра и воронки. В собственно подталампческой области располагается вентро-медиальное гипоталамическое, дорсо-меднальное гипоталамическое, дугообразное, дорсальное гипоталамическое и заднее перивентрикулярное ядра. Ядра промежуточной группы прилежат к углублению воронки III желудочка. К этим ядрам подходят многочисленные сосуды, проникающие в вещество мозга в области заднего продырявленного вещества. Вокруг нейронов формируются капиллярные сплетения. Установлено, что в ядрах промежуточной группы осуществляется анализ химического состава крови и цереброспинальной жидкости. Следовательно, их нейроны обладают хеморецепторными свойствами и в ответ на поступающую информацию о химическом составе крови и цереброспинальной жидкости выделяют релизинг-факторы.
Рилизинг-гормоны, или рилизинг-факторы (release — освобождать), — нейрогормоны, синтезируемые мелкоклеточными ядрами гипоталамуса и стимулирующие (либерины) или угнетающие (статины) выработку и выделение так называемых тропных гормонов гипофиза. Ониобеспечивают взаимодействие высших отделов ЦНС и эндокринной системы. По химической природе рилизинг-гормоны — пептиды. Рилизинг-гормоны выделяются из гипоталамуса в ответ на нервные или химические стимулы и транспортируются с кровью в гипофиз по гипоталамо-гипофизарной портальной системе. Обнаружены 7 стимулирующих (кортиколиберин, тиролиберин, соматолиберин, люлиберин, фоллиберин, пролактолиберин, меланолиберин) и 3 ингибирующих (пролактостатин, меланостатин, соматостатин) секреторную функцию гипофиза рилизинг-гормона. Последние с током крови доставляются в переднюю долю гипофиза (аденогипофиз). Клетки аденогипофиза под воздействием релизинг-факторов (статинов и либеринов) продуцируют тропные гормоны (ТТГ, СТГ, ГТГ, АКТГ, и др.).
3. Задние ядра гипоталамуса представлены задним гипоталамическим ядром. Это ядро имеет непосредственные связи с медиальными ядрами таламуса, с базальными ядрами конечного мозга и с корой полушарий большого мозга. Аксоны клеток заднего гипоталамического ядра заканчиваются на нейронах передних и промежуточных ядер гипоталамуса, следовательно, заднее гипоталамическое ядро в функциональном отношении являются главным среди ядер гипоталамуса. Оно выполняет роль интеграционного центра подталамической области промежуточного мозга. При его поражении у больных развиваются симптомы функциональных нарушений ядер передней и промежуточной групп (нарушение продукции АДГ и тропных гормонов). Кроме названного ядра к задним ядрам гипоталамуса также иногда относят медиальные и латеральные ядра сосцевидного тела.
Учитывая, что гипоталамус координирует нервную и гуморальную регуляцию деятельности всех внутренних органов, его считают высшим центром вегетативных функций организма. В ядрах гипоталамуса осуществляется регуляция сердечно-сосудистой деятельности, температуры тела, различных видов обмена: водного, жирового, углеводного и т.д., выделения слюны, желудочного и кишечного соков, мочи, пота и др.
В свете современных представлений о строении центральной нервной системы указанные высшие центры вегетативных функций находятся под контролем коры полушарий большого мозга.
Мамиллярный комплекс гипоталамуса содержит системы толстых миелинизированных волокон, а также латеральное и медиальное мамиллярные и премамиллярные ядра. Их афференты прежде всего представлены аксонами нейронов гиппокампа, входящими в состав волокон свода. Кроме того, подходят афференты из передней спайки, черной субстанции, ретикулярной формации, коры больших полушарий и мозжечка. Полагают, что кортико- и мозжечково-мамиллярные пути оказывают тормозное влияние на клетки мамиллярного комплекса, а через него и на остальные структуры лимбической системы.
Эпиталамус. Эпиталамическая область расположена дорсально по отношению к каудальным отделам зрительного бугра и занимает относительно небольшой объем. В ее состав входит треугольник поводков, образованный как расширение каудальной части мозговых полосок таламуса и расположенных в его основании ядер поводков. Треугольники соединены комиссурой поводков, в глубине которой проходит задняя комиссура. На поводках – парных тяжах, начинающихся от треугольника, подвешено непарное шишковидное тело, или эпифиз – коническое образование длиной около 6 мм.
Ядра поводков сформированы двумя клеточными группами – медиальными и латеральными ядрами. Афферентами медиального ядра являются волокна мозговых полосок, проводящие импульсацию от лимбических образований конечного мозга (области перегородок, гиппокампа, миндалины), а также от медиального ядра, бледного шара и гипоталамуса. Латеральное ядро получает входы от латеральной преоптической области, внутреннего сегмента бледного шара и медиального ядра. Эфференты медиального ядра, адресованные интерпедункулярному ядру среднего мозга, формируют отогнутый пучок. Эфференты латерального ядра поводков следуют в составе этого же пути, проходят межножковое ядро без переключений и адресуются компактной части черной субстанции, центральному серому веществу среднего мозга и ретикулярным ядрам среднего мозга.
Эпифиз находится посередине под утолщенной задней частью мозолистого тела и располагается в неглубокой борозде, отделяющей друг от друга верхние холмики крыши среднего мозга. Снаружи эпифиз покрыт соединительнотканной капсулой, содержащей большое количество кровеносных сосудов. От капсулы внутрь органа проникают соединительнотканные трабекулы, подразделяющие паренхиму эпифиза на дольки.
Эпифиз является железой внутренней секреции (пинеальная железа) и состоит из глиальных элементов и особых клеток пинеалоцитов. Он иннервируется ядрами поводков, к нему подходят также волокна мозговых полосок задней комиссуры и проекции верхнего шейного симпатического ганглия. Аксоны, входящие в железу, ветвятся среди пинеалоцитов, обеспечивая регуляцию их активности. К числу биологически активных веществ, вырабатываемых эпифизом, относятся мелатонин и вещества, играющие важную роль в регуляции процессов развития, в частности, полового созревания и деятельности надпочечников.
В шишковидном теле у взрослых людей, особенно в старческом возрасте, нередко встречаются причудливой формы отложения, которые придают эпифизу определенное сходство с еловой шишкой, чем и объясняется его название.
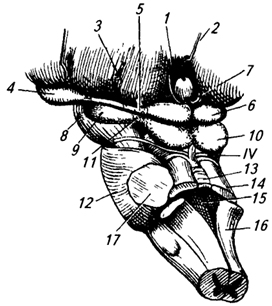
Метаталамус представлен латеральным и медиальным коленчатыми телами – парными образованиями. Они имеют продолговато-овальную форму и соединяется с холмиками крыши среднего мозга при помощи ручек верхнего и нижнего холмиков.
Рис.4. Ствол мозга сбоку и сверху (мозжечок удален):
Латеральное коленчатое тело находится возле нижнебоковой поверхности таламуса, сбоку от его подушки. Его легко можно обнаружить, следуя по ходу зрительного тракта, волокна которого направляются к латеральному коленчатому телу. Несколько кнутри и сзади от латерального коленчатого тела, под подушкой, находится медиальное коленчатое тело, на клетках ядра которого заканчиваются волокна латеральной (слуховой) петли.
Метаталамус состоит из серого вещества. Латеральное коленчатое тело, правое и левое, является подкорковым центром зрения. К нейронам его ядра подходят нервные волокна зрительного тракта (от сетчатки глаза). Аксоны этих нейронов идут в зрительную зону коры. Медиальные коленчатые тела являются подкорковыми центрами слуха.
Третий мозговой желудочек. Полостью промежуточного мозга является третий желудочек. Он представляет собой сагитальную щель, расположенную в срединной плоскости. Его ширина 4-5 мм, длина в верхнем отделе около 25 мм, максимальная высота также 25 мм. Сзади в третий желудочек открывается мозговой водопровод. По бокам своей передней части III желудочек сообщается правым и левым межжелудочковыми отверстиями с боковыми желудочками, лежащими внутри полушарий. Спереди III желудочек ограничен тонкой пластинкой серого вещества – конечной пластинкой, которая представляет собой самую переднюю часть первоначальной стенки мозга, оставшейся посередине между двумя сильно выросшими полушариями. Соединяя оба полушария конечного мозга, эта пластинка и сама принадлежит ему. Непосредственно над ней располагается соединительный пучок волокон, идущих из одного полушария в другое в поперечном направлении; эти волокна связывают участки полушарий, имеющие отношение к обонятельным нервам. Это – передняя комиссура. Ниже конечной пластинки полость III желудочка ограничена перекрестом зрительных нервов.
Боковые стенки III желудочка образованы медиальными сторонами зрительных бугров. На этих стенках проходит продольное углубление – подбугровая борозда. Назад она ведет к водопроводу мозга, вперед – к межжелудочковым отверстиям. Дно III желудочка построено из следующих образований (спереди назад): перекреста зрительных нервов, воронки, серого бугра, сосцевидных тел и заднего продырявленного пространства. Крышу образует эпендема, входящая в состав сосудистых сплетений III и бокового желудочков. Над ней расположен свод и мозолистое тело.
Дата добавления: 2017-03-12 ; просмотров: 3489 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
 | 6984 —
| 6984 —  или читать все.
или читать все.