Синтетическая теория эволюции. Элементарные факторы эволюции. Исследования С.С. Четверикова. Роль эволюционной теории в формировании современной естественнонаучной картины мира.
Когда мы говорим от эволюции, то это понятие у нас всегда ассоциируется с именем Чарлза Дарвина. Жил ученый в 19 в, а споры по поводу эволюции ( в частности, эволюции человека) ведутся по сей день.
Кроме того, генетика, зародившаяся тоже примерно в 19 в, сейчас продвигается вперед просто семимильными шагами. Поэтому, логично было бы объединить имеющиеся знания и накопленный фактический материал в новую теорию эволюции — более современную.
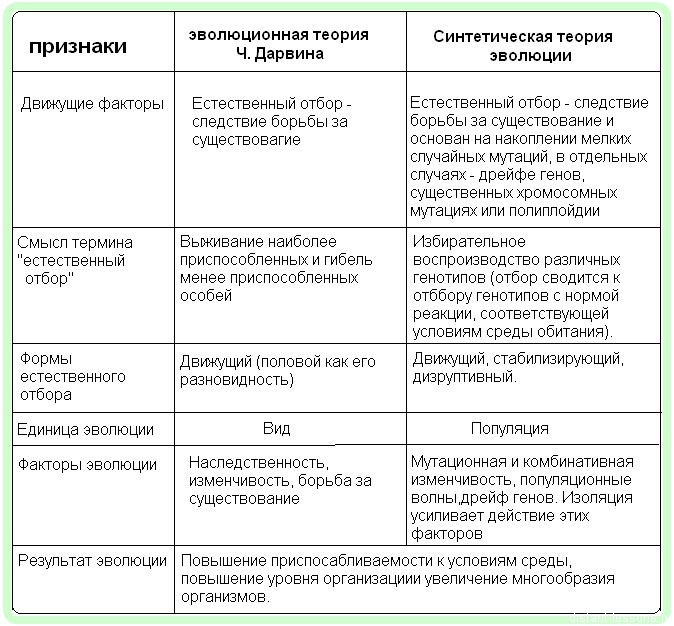
Синтетическая теория эволюции — современный дарвинизм — возникла в начале 40-х годов XX в. Она представляет собой учение об эволюции органического мира, разработанное на основе данных современной генетики, экологии и классического дарвинизма. Термин «синтетическая» идет от названия книги известного английского эволюциониста Дж. Хаксли «Эволюция: современный синтез» (1942). В разработку синтетической теории эволюции внесли вклад многие ученые.
Как и в классической, в синтетической теории эволюции были выдвинуты основные положения:
- Основа эволюции — мутации, причем именно генные, т.к. они должны передаваться по наследству;
- как и в классической теории, в синтетической теории эволюции основной движущий фактор — это естественный отбор ;
- элементарная единица эволюции — популяция;
- эволюция — длительный процесс — смена одной популяции за другой приводит, в конце концов, к образованию вида или нескольких видов (дивергентность);
- вид — это замкнутое образование, при этом наблюдается поток генов — особи мигрируют из одной популяции в другую;
- макроэволюция — результат микроэволюции, при этом все закономерности микроэволюции (на уровне вида) переходят и на более высокий уровень .
Человек, которые впервые объединил знания генетики и эволюционную теорию Дарвиина, — С.С. Четвериков.

Как и Дарвин, Четвериков понимал важность фактического материала — он экспериментально изучал наследование свойств у популяций животных в естественных условиях.
9 лет он работал научным руководителем отдела генетики Института экспериментальной биологии — одного из крупнейших институтов того времени.
В своих работах он показал, что между данными генетики и эволюционной теорией нет никакого противоречия. Напротив, данные генетики должны быть положены в основу учения об изменчивости и стать ключом к пониманию процесса эволюции. Четвериков доказал, что мутации в природных популяциях животных не исчезают, могут накапливаться в скрытом (гетерозиготном) состоянии и давать материал для изменчивости и естественного отбора.
Но, как известно из истории российской (советской) генетики, легкой жизнь наших ученых не была. В конце 30-х гг его осудили и сослали на 6 лет. После ссылки он продолжил свою работу, но уже не в столице, а в городе Горьком… но так же заведующим кафедры генетики Горьковского университета.
Именно он считается основоположником синтетической теории эволюции, ее еще иногда называют «неодарвинизмом», отдавая дань уважения человеку, который все-таки первым выстроил имеющиеся факты в четкую логически обоснованную цепочку теории.
Синтетическая теория эволюции.
Синтетическая теория эволюции (современный дарвинизм) возникла в начале 40-х годов XX в. Она представляет собой учение об эволюции органического мира, разработанное на основе данных современной генетики, экологии и классического дарвинизма. В ней элементарной единицей эволюции служит популяция, поскольку именно в ее рамках происходят наследственные изменения генофонда. Механизм эволюции рассматривается как состоящий из двух частей: случайные мутации на генетическом уровне и наследование наиболее удачных с точки зрения приспособления к окружающей среде мутаций, так как их носители выживают и оставляют потомство.
Становление теории началось с созданной в 1926 году С.С. Четвериковым популяционной генетики. Из его работ стало ясно, что отбору подвергаются не отдельные признаки и отдельные особи, а генотип всей популяции. Через фенотипические признаки отдельных особей осуществляется отбор генотипов популяции, ведущий к распространению полезных изменений. Затем в создание новой теории включились около 50 ученых из восьми стран, их коллективными трудами и была создана СТЭ. Термин «синтетическая» идет от названия книги известного английского эволюциониста Дж. Хаксли «Эволюция: современный синтез» (1942).
Структурно СТЭ состоит из теорий микро- и макроэволюции:
1) Теория микроэволюции изучает необратимые преобразования генетико-экологической структуры популяции, которые могут привести к формированию нового вида. Реально вид существует в виде популяций. Именно популяция является элементарной единицей эволюции.
2) Теория макроэволюции изучает происхождение надвидовых таксонов (семейств, отрядов, классов и т.д.), основные направления и закономерности развития жизни на Земле в целом, включая возникновение жизни и происхождение человека как биологического вида.
Изменения, которые изучаются в рамках микроэволюции, доступны непосредственному наблюдению, тогда как макроэволюция происходит на протяжении длительного исторического периода времени и поэтому ее процесс может быть только реконструирован задним числом. Но макро- и микроэволюция происходят в конечном итоге под воздействием изменений в окружающей среде.
Основные положения синтетической теории эволюции:
Материалом для эволюции служат наследственные изменения — мутации (как правило, генные) и их комбинации.
Основным движущим фактором эволюции является естественный отбор, возникающий на основе борьбы за существование.
Наименьшей единицей эволюции является популяция.
Эволюция носит в большинстве случаев дивергентный характер, т. е. один таксон может стать предком нескольких дочерних таксонов.
Эволюция носит постепенный и длительный характер. Видообразование как этап эволюционного процесса представляет собой последовательную смену одной временной популяции чередой последующих временных популяций.
Вид состоит из множества соподчиненных, морфологически, физиологически, экологически, биохимически и генетически отличных, но репродуктивно не изолированных единиц — подвидов и популяций.
Вид существует как целостное и замкнутое образование. Целостность вида поддерживается миграциями особей из одной популяции в другую, при которых наблюдается обмен аллелями («поток генов»),
Макроэволюция на более высоком уровне, чем вид (род, семейство, отряд, класс и др.), идет путем микроэволюции. Согласно синтетической теории эволюции, не существует закономерностей макроэволюции, отличных от микроэволюции. Иными словами, для эволюции групп видов живых организмов характерны те же предпосылки и движущие силы, что и для микроэволюции.
Любой реальный (а не сборный) таксон имеет монофилетическое происхождение.
Эволюция имеет ненаправленный характер, т. е. не идет в направлении какой-либо конечной цели.
Н.В. Тимофеев-Ресовский сформулировал положение об элементарных явлениях и факторах эволюции: 1) популяция -элементарная эволюционная структура; 2) изменение генотипического состава популяции — элементарное эволюционное явление; 3) генофонд популяции — элементарный эволюционный материал; 4) элементарные эволюционные факторы — мутационный процесс, «волны жизни», изоляция, естественный отбор.
Оказалось, что популяция в качестве элементарной структуры должна быть способной изменяться с течением времени и должна реально существовать в природных условиях. Тогда ее определение таково: популяция — это совокупность особей данного вида, занимающих территорию внутри ареала вида, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.
В свою очередь элементарным эволюционным явлением считаются наследственные изменения популяций, в результате спонтанных мутаций, представляющих собой гетерогенную смесь различных генотипов. Изменения эти тем отчетливее, чем более интенсивно и длительно воздействие факторов, их вызывающих. В результате происходит изменение генофонда, или генотипического состава популяции.
Еще одно требование к популяциям, выступающим в качестве единиц эволюции, — способность трансформироваться в элементарный эволюционный материал. А это осуществимо при следующих условиях: 1) у всех особей, составляющих популяцию, должны происходить наследственные изменения материальных единиц; 2) эти изменения должны затрагивать все свойства особей, вызывая их отклонения от исходных; 3) они должны затрагивать биологически важные свойства особей; 4) изменения эти должны быть четко выражены у популяций, обитающих в природных условиях; 5) часть таких изменений должна «выходить» на историческую арену эволюции, участвуя в образовании таксонов низшего ранга; 6) скрещивающиеся таксоны должны различаться наборами и комбинациями элементарных единиц наследственной изменчивости.
Согласно постулатам СТЭ, требованиям элементарного эволюционного материала удовлетворяют различного рода мутации. К их числу относят генные, хромосомные, геномные мутации. Чтобы мутации служили материалом эволюции, необходимы: достаточная частота возникновения мутаций, четкость в проявлении мутантных признаков и четко выраженная биологическая значимость этих признаков, генетические различия между природными таксонами.
Не менее важны и так называемые элементарные эволюционные факторы, воздействующие на количественные соотношения генов конкретной популяции. Такого рода факторы должны удовлетворять следующим требованиям: 1) быть поставщиком элементарного эволюционного материала, необходимого для проявления элементарного эволюционного явления — изменения генотипического состава популяции; 2) расчленять исходную популяцию на две или несколько, разделенные различными изоляционными барьерами; 3) создавать внутрипопуляционные барьеры; 4) вызывать адаптивные изменения.
Первый фактор, удовлетворяющий вышеназванным требованиям, это мутационный процесс, одновременно являющийся и поставщиком элементарного эволюционного материала. Но сам по себе этот фактор не способен оказывать направляющее воздействие на эволюционный процесс. Для этого нужен второй фактор — популяционные волны, или «волны жизни», — количественные колебания в численности популяций под воздействием различных причин — сезонной периодики, климатических, природно-катастрофических и пр.
Эволюционная роль «волн жизни» проявляется в двух планах. Во-первых, в изменении частот генов в популяциях, приводящем к снижению наследственной изменчивости. Процесс этот, названный американским генетиком С. Райтом «дрейфом генов», а Н.П. Дубининым — «генетико-автоматическим процессом», всегда имеет место при резком снижении численности популяции. Генотипически это сопровождается увеличением гомозиготности, что связано с увеличением числа близкородственных скрещиваний. Другое проявление «волн жизни» сводится к изменениям в концентрации различных мутаций, а также к уменьшению разнообразия генотипов, содержащихся в популяции. А это в свою очередь может привести к изменениям направленности и интенсивности действия отбора.
Третий элементарный эволюционный фактор — это изоляция. Нарушая свободное скрещивание, изоляция закрепляет возникшие как случайно, так и под действием отбора различия в наборах и численности генотипов в изолированных частях популяции. Различают два типа изоляции: территориально-механическую, или пространственно-географическую, и биологическую, или репродуктивную. Смысл первой ясен из названия. Биологическая же изоляция имеет пять форм: этологическую (различия в поведении особей), экологическую (различия в предпочтении разных мест обитания), сезонную (различия в сроках размножения), морфологическую (различия в размерах, структуре как всего тела организма, так и отдельных его органов), генетическую (различия наследственного аппарата, приводящие к несовместимости половых клеток). Общим итогом изоляции является возникновение независимых генофондов двух популяций, которые в итоге могут трансформироваться в самостоятельные виды.
Четвертый элементарный эволюционный фактор — естественный отбор. Его генетическая сущность — дифференцированное (неслучайное) сохранение в популяции определенных генотипов и избирательное их участие в передаче генов следующему поколению. Здесь важно подчеркнуть, что естественный отбор воздействует не на отдельный фенотипический признак, не на отдельный ген, то есть молекулярно-генетическую систему. Его роль разыгрывается на уровне фенотипа, то есть целостной живой системы — организма, сформированного в результате взаимодействия с генотипом, имеющим определенную норму реакции.
В настоящее время известны три формы отбора. Это движущий отбор, при котором в результате новых мутаций или перекомбинаций уже имеющихся генотипов или при изменении условий среды в популяции возникают новые генотипы с селективными свойствами. Тогда может возникнуть новый вектор, или направленность, отбора. Под контролем такого отбора генофонд популяции изменяется как единое целое, то есть отсутствует дивергенция дочерних форм.
Второй вид отбора получил название стабилизирующего. Его роль сводится к тому, что в конкретных условиях на основе разных генотипов в популяции становится преобладающим оптимальный для этих условий фенотип. При длительной неизменности таких условий стабилизирующий отбор как бы охраняет ставший устойчивым фенотип от давления любой фенотипической изменчивости.
Третья форма отбора называется дизруптивной. Ее роль в том, чтобы внутри популяции могли возникнуть отчетливо различающиеся формы. При снижении возможности скрещивания между такими популяциями, например, в условиях изоляции, может происходить их дальнейшее расхождение, вплоть до образования новых видов.
Синтетическая теория эволюции вскрыла глубинные механизмы эволюционного процесса, накопила множество новых фактов и доказательств эволюции живых организмов, объединила данные многих биологических наук. Тем не менее синтетическая теория эволюции (или неодарвинизм) находится в русле тех идей и направлений, которые были заложены Ч. Дарвином.
Синтетическая теория эволюции
Предполагается, что трудности, с которыми сталкивалась классическая теория эволюции (см. п. 5.5.2), в частности при объяснении явления наследственности, были преодолены путем синтеза эволюционной теории Ч. Дарвина и генетики Г. Менделя.
Синтетическая теория эволюции (СТЭ) — это современная эволюционная теория, которая является синтезом различных дисциплин, прежде всего генетики и дарвинизма, а также палеонтологии, систематики, молекулярной биологии и др.
Синтетическая теория в ее нынешнем виде образовалась в результате переосмысления ряда положений классического дарвинизма с позиций генетики начала XX в.
После переоткрытия законов Г. Менделя (в 1901 г.), доказательства дискретной природы наследственности и, особенно, после создания теоретической популяционной генетики трудами Р. Фишера, Д.Б.С. Холдейна-младшего и С. Райта учение Ч. Дарвина приобрело прочный генетический фундамент.
Статья С.С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» [103] стала ядром будущей и основой для дальнейшего синтеза дарвинизма и генетики. В этой статье показана совместимость принципов генетики с теорией естественного отбора и заложены основы эволюционной генетики.
В работах Дж. Холдейна, Н.В. Тимофеева-Ресовского и Ф.Г. Добр- жанского идеи, выраженные С.С. Четвериковым, распространились на Запад, где почти одновременно Р. Фишер высказал взгляды об эволюции доминантности.
Толчок к развитию СТЭ дала гипотеза о рецессивности новых генов. Гипотеза предполагала, что в каждой воспроизводящейся группе организмов во время созревания гамет в результате ошибок при репликации ДНК постоянно возникают мутации — новые варианты генов.
Влияние генов на строение и функции организма таково, что каждый ген участвует в определении нескольких признаков. С другой стороны, каждый признак зависит от многих генов. Генетики называют это явление генетической полимерией признаков.
Английский генетик Р. Фишер в 1930 г. высказал идею о том, что полимерия отражает взаимодействие генов, в связи с чем внешнее проявление каждого гена зависит от его генетического окружения. Поэтому рекомбинация, порождая все новые генные сочетания, в конце концов создает для данной мутации такое генное окружение, которое позволяет мутации проявиться в фенотипе особи-носителя.
Так мутация попадает под действие естественного отбора, который уничтожает сочетания генов, затрудняющие жизнь и размножение организмов в данной среде, и сохраняет нейтральные и выгодные сочетания. Причем отбираются прежде всего такие генные комбинации, которые способствуют благоприятному и одновременно устойчивому фенотипическому выражению изначально малозаметных мутаций, за счет чего эти мутантные гены постепенно становятся доминантными.
Таким образом, сущность синтетической теории составляет преимущественное размножение определенных генотипов и передача их особенностей потомкам. В вопросе об источнике генетического разнообразия синтетическая теория признает главную роль за рекомбинацией генов.
Считают, что эволюционный акт состоялся, когда отбор сохранил генное сочетание, нетипичное для предшествующей истории вида.
Важной предпосылкой для возникновения новой теории эволюции стала книга английского генетика, математика и биохимика
Дж. Б.С. Холдейна-младшего «The causes of evolution» (1932). Холдейн, создавая генетику индивидуального развития, сразу же включил новую науку в решение проблем макроэволюции.
Эволюционные новшества часто возникают на основе неотении (сохранение ювенильных признаков у взрослого организма). Неотенией Дж. Холдейн объяснял происхождение человека («голая обезьяна»), эволюцию других крупных таксонов. В 1933 г. Н.К. Кольцов, учитель С.С. Четверикова, показал, что неотения в животном царстве широко распространена и играет важную роль в прогрессивной эволюции. Она ведет к морфологическому упрощению, но при этом сохраняется богатство генотипа.
Практически во всех историко-научных моделях 1937 г. был назван годом возникновения СТЭ. Именно в этом году появилась книга русско-американского генетика и натуралиста Ф.Г. Добржанского. Двойная специализация Ф.Г. Добржанского позволила ему первому перебросить твердый мост от лагеря экспериментальных биологов к лагерю натуралистов.
Впервые было сформулировано важнейшее понятие об «изолирующих механизмах эволюции» — репродуктивных барьерах, которые отделяют генофонд одного вида от генофондов других видов. Ф. Г. Добржанский также внедрил в натуралистический материал «эффект С. Райта», полагая, что микрогеографические расы возникают под воздействием случайных изменений частот генов в малых изолятах, т.е. адаптивно-нейтральным путем.
В англоязычной литературе среди создателей СТЭ чаще всего называют имена Ф.Г. Добржанского, Дж. Хаксли, Э. Майра, Б. Ренша, Дж. Стеббинса. Это, конечно, не полный список. Из русских ученых, по меньшей мере, следовало бы назвать И.И. Шмальгаузена, Н.В. Тимофеева-Ресовского, Г.Ф. Гаузе, Н.П. Дубинина, А.Л. Тахтаджяна; из британских ученых — Дж.Б.С. Холдейна-младшего, Д. Лэка, К. Уод- дингтона, Г. де-Бира; из немецких ученых — Э. Баура, В. Циммермана, В. Людвига, Г. Хеберера и др. [74].
В результате в 1930—1940 гг. была создана синтетическая теория эволюции, не только ставшая ядром популяционной генетики, но и позволившая сформулировать единую систему всего современного биологического знания.
В отличие от классической эволюционной концепции Ч. Дарвина, рассматривающей в качестве единицы эволюции вид, СТЭ утверждает, что элементарной эволюционной структурой является популяция.
Полагали, что именно популяция обладает теми свойствами самоорганизующейся целостной системы, которые необходимы для наследственных изменений.
Устойчивое изменение генотипа популяции рассматривается в качестве элементарного явления эволюционного процесса. Единицей наследственности является ген — участок молекулы ДНК, отвечающий за развитие определенных признаков организма.
Основной механизм эволюционного процесса — отбор организмов с полезными, выгодными для приспособления к среде обитания мутациями.
Наследственные изменения происходят под действием ряда эволюционных факторов:
- • мутационный процесс — мутационные изменения, поставляющие материал для эволюции;
- • популяционные волны- колебания численности популяции вокруг некоторого среднего уровня;
- • изоляция — обособление популяции для закрепления нового признака;
- • естественный отбор — ведущий фактор эволюции — выживание наиболее приспособленных особей и рождение ими здорового потомства.
Мутации — это изменение наследственных свойств организмов внутри популяции, возникающее естественным или искусственным путем и поставляющее основной материал для эволюции. Как уже упоминалось, мутагенами являются температурный режим, действие отравляющих веществ, радиация, особенности питания и т.п. [78].
После открытия двойной спирали ДНК (1953) мутацию стали трактовать в духе моргановской хромосомной теории: в ней видели изменение в тексте ДНК — в структуре нуклеиновой кислоты в пределах локуса — или в строении хромосом. Мутации стали разделять на генные (точковые), хромосомные и геномные. Казалось, что к этим трем типам мутаций сводится любое наследственное изменение. В связи с таким ограничением и стала возможной разработка генетико-популяционной модели эволюции в СТЭ.
Вместе с постулатом, что единственным источником эволюции являются мутации, в СТЭ прочно утвердилось представление об однозначном соответствии мутации (гена) и признака, о том, что возникновение нового устойчивого фенотипа является автоматическим следствием проявления мутации. С этих позиций эволюция предстает как результат сортировки и накопления естественным отбором серии мутаций
Современная молекулярная биология к числу наиболее опасных мутагенов относит вирусы.
Мутации появляются случайно, большинство из них либо опасны, либо вредны. Вредные мутации часто вызывают гибель организма, причем, как правило, на достаточно ранних этапах онтогенеза; вредные мутации, не приведшие к летальному исходу, элиминируются в ходе естественного отбора.
Благоприятные мутации крайне редки, но именно они дают организму эволюционное преимущество. Случайные благоприятные мутации постепенно накапливаются в популяции, закрепляются в ряде поколений и способствуют эволюции вида.
Популярные волны, или волны численности, которые иногда называют «волнами жизни», определяют колебания численности популяции вокруг некоторой средней величины. Исследования показали, что наиболее благоприятны для появления новых свойств и возникновения новых видов популяции среднего размера.
Изоляция — еще один фактор эволюционного процесса, необходимый для того, чтобы популяция не могла скрещиваться с другими группами организмов и обмениваться с ними генетической информацией.
Целесообразность в живой природе является следствием естественного отбора, который выступает движущей силой и ведущим фактором эволюции. Отбор действует на всех этапах развития живого организма, ему подвергаются все без исключения свойства. В классической теории эволюции естественный отбор определялся как процесс выживания наиболее приспособленных организмов.
Современная эволюционная биология делает акцент на другой стороне этого явления. Естественный отбор теперь понимается как устранение от размножения тех особей, которые менее приспособлены к условиям внешней среды.
Перечисленные факторы эволюции действуют как на микроэво- люционном (эволюционные изменения в популяции за небольшой период времени), так и на макроэволюционном уровне (совокупность эволюционных изменений на протяжении длительного времени, приводящих к возникновению новых надвидовых форм организации живого).
Важной составной частью СТЭ являются концепции микро- и макроэволюции.
Под микроэволюцией понимают совокупность эволюционных процессов, протекающих в популяциях, приводящих к изменениям генофонда этих популяций и образованию новых видов.
Считается, что микроэволюция протекает на основе мутационной изменчивости под контролем естественного отбора. Мутации — единственный источник появления качественно новых признаков, а естественный отбор — единственный творческий фактор микроэволюции.
На характер процессов микроэволюции оказывают влияние колебания численности популяций («волны жизни»), обмен генетической информацией между ними, их изоляция и дрейф генов. Микроэволюция ведет либо к изменению всего генофонда биологического вида как целого, либо к обособлению от родительского вида в качестве новых форм.
Пор макроэволюцией понимают эволюционные преобразования, ведущие к формированию таксонов более высокого ранга, чем вид (родов, отрядов, классов).
Считается, что макроэволюция не имеет специфических механизмов и осуществляется только посредством процессов микроэволюции, будучи их интегрированным выражением. Накапливаясь, микроэво- люционные процессы выражаются внешне в макроэволюционных явлениях, т.е. макроэволюция представляет собой обобщенную картину эволюционных изменений. Поэтому на уровне макроэволюции обнаруживаются общие тенденции, направления и закономерности эволюции живой природы, которые не поддаются наблюдению на уровне микроэволюции. (Об эволюции не по Ч. Дарвину см. гл. 16.)
Основные положения синтетической теории эволюции
Синтетическая теория эволюции — современный дарвинизм — возникла в начале 40-х годов XX в. Она представляет собой учение об эволюции органического мира, разработанное на основе данных современной генетики, экологии и классического дарвинизма. Термин «синтетическая» идет от названия книги известного английского эволюциониста Дж. Хаксли «Эволюция: современный синтез» (1942). В разработку синтетической теории эволюции внесли вклад многие ученые.
Основные положения синтетической теории эволюции в общих чертах можно выразить следующим образом:
1. Материалом для эволюции служат наследственные изменения — мутации (как правило, генные) и их комбинации.
2. Основным движущим фактором эволюции является естественный отбор, возникающий на основе борьбы за существование.
3. Наименьшей единицей эволюции является популяция.
4. Эволюция носит в большинстве случаев дивергентный характер, т. е. один таксон может стать предком нескольких дочерних таксонов.
5. Эволюция носит постепенный и длительный характер. Видообразование как этап эволюционного процесса представляет собой последовательную смену одной временной популяции чередой последующих временных популяций.
6. Вид состоит из множества соподчиненных, морфологически, физиологически, экологически, биохимически и генетически отличных, но репродуктивно не изолированных единиц — подвидов и популяций.
7. Вид существует как целостное и замкнутое образование. Целостность вида поддерживается миграциями особей из одной популяции в другую, при которых наблюдается обмен аллелями («поток генов»),
8. Макроэволюция на более высоком уровне, чем вид (род, семейство, отряд, класс и др.), идет путем микроэволюции. Согласно синтетической теории эволюции, не существует закономерностей макроэволюции, отличных от микроэволюции. Иными словами, для эволюции групп видов живых организмов характерны те же предпосылки и движущие силы, что и для микроэволюции.
9. Любой реальный (а не сборный) таксон имеет монофилети-ческое происхождение.
10. Эволюция имеет ненаправленный характер, т. е. не идет в направлении какой-либо конечной цели.
Синтетическая теория эволюции вскрыла глубинные механизмы эволюционного процесса, накопила множество новых фактов и доказательств эволюции живых организмов, объединила данные многих биологических наук. Тем не менее синтетическая теория эволюции (или неодарвинизм) находится в русле тех идей и направлений, которые были заложены Ч. Дарвином.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Только сон приблежает студента к концу лекции. А чужой храп его отдаляет. 8019 —  | 6883 —
| 6883 —  или читать все.
или читать все.
193.124.117.139 © studopedia.ru Не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования. Есть нарушение авторского права? Напишите нам | Обратная связь.
Отключите adBlock!
и обновите страницу (F5)
очень нужно
2.1. Становление и основные положения синтетической теории эволюции
2.1. Становление и основные положения синтетической теории эволюции
Эволюционизм возник как альтернатива учению о неизменности видов. Вопросы, связанные с возникновением и развитием жизни, прошли через всю интеллектуальную историю человечества. Количество литературы, посвященной истории эволюционных учений, огромно (Завадский К. М., Колчинский Э. И., 1977; Колчинский Э. И., 2002).
Термин «эволюция» впервые был предложен швейцарским натуралистом Ш. Боннэ (1720–1793), который рассматривал ее как процесс развертывания возможностей, заложенных в материю Творцом. В Новое время первые, весьма несовершенные, концепции эволюции мы находим у французского натуралиста Ж. Бюффона (1707–1788) и деда Ч. Дарвина – Э. Дарвина (1731–1807).
Большую известность (правда, уже после смерти автора) приобрела теория французского естествоиспытателя Ж. Б. Ламарка (1744–1829). Свое эволюционное учение он изложил в знаменитой книге «Философия зоологии» (1809), где развивал взгляды о постепенном повышении уровня организации живых существ от простейших до человека.
Ж. Б. Ламарк выделил три основных фактора эволюции: упражнение органов, наследование благоприобретенных признаков, внутреннее стремление организмов к прогрессу. Хотя наследование благоприобретенных признаков во времена Ламарка считалось само собой разумеющимся, его теория не получила признания среди современников. Многие обоснования Ж. Б. Ламарка были неубедительны и явно натянуты. Большинство биологов того времени, в том числе и крупнейший авторитет – французский палеонтолог Ж. Кювье (1769–1832), стояли на позициях креационизма и неизменности видов.
Следует заметить, что эволюция не обязательно должна противопоставляться креационизму. Креационизм постулирует только акт творения жизни, но может допускать ее последующую эволюцию, что мы видим уже у Ш. Боннэ.
В трудах последователей Ж. Б. Ламарка его теория получила развитие, став влиятельным течением эволюционизма. Однако адекватному пониманию его идей часто мешал идеологический фактор. Только сейчас мы начинаем постигать их значение. Несмотря на ошибки, объяснимые уровнем развития биологии того времени, мысли Ж. Б. Ламарка оказались исключительно глубоки.
Из других эволюционистов того времени нельзя не отметить работы французского естествоиспытателя Э. Жоффруа Сент-Илера (1772–1844).
Однако крупнейшее событие в науке XIX в. – это эволюционная теория Ч. Дарвина (1809–1882). Суть теории Ч. Дарвин изложил в своей эпохальной книге «Происхождение видов», напечатанной в 1859 г. С этой даты многие историки науки ведут отсчет современной биологии. Книга произвела настоящую революцию и в научном мировоззрении, и в умах людей того времени. Дарвинизм быстро стал господствующим течением эволюционизма.
Основными факторами эволюции Ч. Дарвин выдвинул изменчивость, наследственность и отбор. Главным фактором у него выступает естественный отбор. О механизмах наследственности и изменчивости Ч. Дарвин, как и Ж. Б. Ламарк, имел самые смутные представления. Однако, вне зависимости от механизма, изменчивость у него не была привязана к «желаниям» организма или к «высшим силам». Непрерывный ряд мелких изменений от первоначальной формы и составляет материал эволюции. Ч. Дарвин снял мистический ореол таинственности с эволюционного процесса.
Существенно потеснив креационизм в общественном сознании, дарвинизм не остался единственной эволюционной теорией. Дав мощный толчок эволюционному подходу в науке, он одновременно породил альтернативные версии. Одним из самых известных критиков дарвинизма XIX в. был русский зоолог Н. Я. Данилевский (1822–1895), который провел скрупулезный анализ работ Ч. Дарвина.
Начало XX в. отмечено периодом долгой конфронтации дарвинизма с нарождающейся генетикой. Это породило обилие антидарвиновских концепций, предрекающих «скорую смерть» теории Ч. Дарвина. Наиболее уязвимой стороной дарвинизма в то время считалась природа наследственности. Поэтому рождение синтетической теории эволюции (СТЭ), которая базировалась на синтезе популяционной генетики и дарвинизма, стали рассматривать как вторую революцию в истории эволюционной биологии.
Теоретические положения популяционной генетики были разработаны С. Райтом (1889–1988), Р. Фишером (1890–1962), Дж. Холдейном (1860–1936) в 1930–1931 гг. Они изложены в классических работах «Генетическая теория естественного отбора» (Fisher R., 1930) и «Факторы эволюции» (Haldane J., 1932).
Контуры целостной эволюционной теории впервые были очерчены в работе Ф. Добжанского «Генетика и происхождение видов» (Dobzhansky Th., 1937). Часто называют шесть главных «архитекторов» СТЭ: Т. Добжанский (1900–1975), Э. Майр (1904–2005), Б. Ренш (1900–1990), Н. В. Тимофеев-Ресовский (1900–1981), Дж. Симпсон (1902–1984), Дж. Хаксли (1887–1975). Однако на самом деле в создание нового направления весьма весомый вклад внесли более тридцати биологов разных специальностей, что отмечают в своих работах и сами «отцы-основатели». Можно вспомнить таких известных ученых, как К. Дарлингтон (1903–1981), Англия, Г. Хеберер (1901–1973), Германия, Дж. Стеббинс (1906–2000), США. Достойное место в этом ряду занимает наш соотечественник И. И. Шмальгаузен (1884–1963). Сам термин «синтетическая теория эволюции» вошел в употребление после выхода книги Дж. Хаксли «Эволюция. Современный синтез» (Huxley J., 1942).
С точки зрения СТЭ, эволюция – это процесс постепенного ненаправленного изменения частот и видов аллелей во многих локусах.
Элементарной единицей эволюционного процесса в СТЭ была признана популяция – изолированная группа особей одного вида, связанная общностью территории и происхождения. Этот термин был предложен В. Иоганнсеном в 1908 г. Совокупность аллелей популяции называется генофондом, а процессы, изменяющие генофонд, получили название элементарных эволюционных факторов. По поводу числа факторов среди эволюционистов нет единодушия. Наиболее часто в СТЭ выделяют как эволюционные факторы мутационный процесс, поток генов, дрейф генов и естественный отбор.
Мутационный процесс – процесс образования новых генетических вариантов. Исключительно важным аспектом СТЭ является положение о случайном, ненаправленном характере мутаций. Поскольку мутации – редкое событие, то они изменяют генофонд чрезвычайно медленно.
Поток генов – обмен генами между разными популяциями. Основной источник потока генов – миграции особей, которые способны изменять генофонд значительно быстрее, чем мутации. Диапазон интенсивности миграций различен. Обмен генов между популяциями может вообще отсутствовать. В таком случае мы наблюдаем изоляцию – важнейший фактор видообразования.
Дрейф генов – случайные изменения частот аллелей в популяции. Это понятие в генетику ввел С. Райт (Wright S., 1931). Оно служит предметом долгих дискуссий. Дрейф генов относится к явлениям, во многом обусловленным ошибкой выборки: чем меньше выборка популяции, тем больше будет ошибка выборки.
Естественный отбор – важнейший фактор эволюции в СТЭ. Кратким и удачным определением отбора может служить определение, данное И. Лернером: «отбор – это дифференциальное воспроизведение генотипов» (Lerner I., 1958). Данное определение подразумевает, что шансы передать свои признаки следующему поколению у разных генотипов не одинаковы. Один из основателей цитогенетики, английский генетик С. Дарлингтон (1903–1981) охарактеризовал естественный отбор как процесс переноса «…с химического уровня мутации на биологический уровень адаптации» (Darlington C., 1958). В СТЭ отбор рассматривается как единственный направляющий эволюционный фактор. Роль естественного отбора – это один из ключевых дискуссионных вопросов на всем протяжении истории эволюционизма после Ч. Дарвина.
Другим ключевым вопросом стал вопрос единства механизмов эволюции. В СТЭ принято различать разные уровни эволюционного процесса.
Микроэволюция – процесс адаптивного изменения популяций до возникновения новых видов. Все вышерассмотренные закономерности динамики популяций под действием эволюционных факторов разработаны именно по отношению к микроэволюции.
Макроэволюция – эволюция надвидовых таксонов. Это один из наименее разработанных разделов СТЭ, можно сказать, ее слабое место. Среди сторонников СТЭ преобладают взгляды о единстве механизмов микро– и макроэволюции, что во многом обусловлено представлениями «отцов-основателей» Дж. Симпсона, Б. Ренша, Дж. Хаксли. Это же относится и к пограничному явлению – видообразованию.
Видообразование – процесс образования новых видов. Оно обычно рассматривается как итог микроэволюции, но ряд эволюционистов предлагают считать видообразование самостоятельным разделом.
Таковы (в самом общем виде) основные положения синтетической теории эволюции. Ее становление, можно сказать, проходило триумфально. Середина XX в. знаменуется безраздельным господством идей СТЭ. Хотя и в это время выдвигались альтернативные теории, в целом ведущие эволюционисты проявляли редкое единодушие. Теоретические затруднения рассматривались как естественные аспекты развития науки.
Затем положение изменилось. Новые факты, порождая новые разногласия, поставили эволюционную биологию на порог третьей научной революции. Чтобы понять суть теоретических разногласий, необходимо кратко познакомиться с историей альтернативных концепций. Они возникли, как уже говорилось, почти одновременно с дарвинизмом.
Синтетическая теория эволюции кратко и понятно. Прав ли Дарвин?
Теория эволюции Дарвина, опубликованная впервые в далеком 1859 году, до сих пор вызывает чувство отторжения и неприязни у многих наших сограждан. Причины этого вполне понятны. Происхождение человека из царства животных, роднящее наш божественный разум с инстинктами обезьян, рефлексами ящериц и даже с химическими реакциями сине-зеленых водорослей, предполагает следствия, несовместимые с утешительными и возвышенными доктринами религий. Поэтому многие никак не могут успокоится и постоянно ищут опровержений теории Чарльза Дарвина, иногда предлагая довольно странные конструкции, демонстрирующие плохое понимание сути вопроса.
Одним из удивительных выводов сторонников креационизма и неизменности живого на нашей планете является заявление о том, что синтетическая теория эволюции опровергает построения Дарвина. Давайте разберемся, что такое эта самая синтетическая теория и правда ли, что в ней содержатся положения, несовместимые с дарвиновским естественным отбором.
Как появилась синтетическая теория эволюции
Бурное развитие науки в конце XIX и начале ХХ века породило несколько важнейших направлений исследования живой природы. Помимо дарвинизма, определившего развитие биологии, в 1865 году были обнародованы законы Менделя. Тогда еще никто не понимал, что монах Грегор Мендель открыты механизмы передачи признаков по наследству, блестяще предсказанный но не обнаруженный Дарвином. О результатах, полученных Менделем вскоре все забыли, кроме селекционеров, работавших с новыми сортами растений, что и неудивительно: по сравнению с общественным резонансом на сенсационные заявления «Происхождения видов», какие-то жалкие эксперименты с горохом не привлекали внимания общественности.
С другой стороны развивалась молекулярная биология. Ученые стремились проникнуть внутрь клетки и понять, как работают ее органеллы. В 1882-м году в свет вышла книга немецкого профессора Вальтера Флемминга «Клеточное вещество, ядро и клеточное деление», в которой были собраны все имеющиеся на тот момент сведения о делении клетки и о хромосомах. Уже к 1902-му году на свет были вновь извлечены законы старика Менделя и стало ясно, что именно хромосомы содержат наследственную информацию организма.
Генетика Менделя изучала внешние проявления наследственности, а молекулярная биология, родившаяся из исследований Флеминга, рассматривала внутренние механизмы передачи признаков потомству. И оба эти направления прекрасно подтверждали предсказания Дарвина о наличии некоего механизма наследственности, присущего живой природе.
В 1937 году вышла книга Ф.Г. Добржанского «Генетика и происхождение видов», показавшая что все эти направления — генетика, молекулярная биология и теория эволюции прекрасно дополняют друг друга, образуя прекрасную и полную общую картину развития жизни на Земле. Так, из объединения нескольких областей знания, возникла принятая в настоящее время синтетическая (объединенная) теория эволюции (далее СТЭ). Не стоит полагать, что она опровергает теорию естественного отбора, напротив, дополняет и подтверждает тезисы, выведенные Дарвином.
Чем синтетическая теория эволюции отличается от дарвинизма
Наука не стоит на месте. Фонд знаний постоянно пополняется, гипотезы строятся, опровергаются, теории получают фактическое подтверждение или меняются под влиянием фактов. Так блестяще подтвердились предположения Дарвина о переходных формах, в изобилии обнаруженных палеонтологами для разных групп животных, в том числе и для человека. Естественно, и наше понимание эволюционных процессов несколько изменилось с течением времени.
Например, СТЭ выделяет популяцию в качестве минимальной эволюционной единицы. Дарвин же рассматривал в качестве таковой вид целиком. Под процессом эволюции современная наука понимает устойчивое изменение генотипа популяции, о чем Дарвин просто не знал в свое время. СТЭ шагнула далеко вперед по сравнению с представлениями Дарвина об эволюции, выделив множество факторов, являющихся движущей силой эволюции. Там, где Дарвин обходился внутри- и межвидовой борьбой за существование и борьбой с неблагоприятными условиями неживой природы, сейчас выделяют мутационные процессы, изоляцию, популяционные волны численности и другие факторы. При этом мутационная и рекомбинационная изменчивость организмов Дарвину не были известны. Он рассматривал процесс накопления изменений без понимания механизмов, его обуславливающих. Так же Дарвину не было известно ни о дрейфе генов, не о «принципе основателя», основной причины формирования нейтральных признаков, которые Дарвин не мог объяснить.
Другими словами, Дарвин был прав во всем, но не знал того, что сейчас написано в каждом учебнике по биологии. Он описывал действие эволюции основываясь на наблюдениях и предположениях, которые блестяще подтвердились в наше время.
В основе синтетической теории эволюции (СТЭ), или эволюционного синтеза, лежит все тот же дарвинизм. Однако он дополнен сведениями из других биологических наук, в первую очередь генетики, а также экологии, молекулярной биологии, систематики и др.
Многие особенности и закономерности эволюционного процесса, описанные Дарвином, не могли быть им объяснены в полной мере из-за недостаточной развитости наук в то время. Это послужило почвой для обоснованной критики эволюционной теории. Например, Дженкин заметил, что любое изменение, возникшее в одной особи постепенно растворится в результате скрещивания и, следовательно, не может закрепиться в результате борьбы за существование (так называемый «кошмар Дженкина»). Дарвин и его последователи не могли привести контраргументы, так как мало себе представляли дискретную природу наследственности, хотя и открытую Менделем в 60-х годах XIX, но признанную в науке лишь в начале XX века.
Открытия в области генетики, молекулярной биологии, исследования популяций и приход к пониманию, как и почему популяция является единицей эволюции, привели к тому, что эволюционная теория уже перестала быть чем-то вроде гипотезы, а была объяснена и во многом доказана. Были более глубоко вскрыты механизмы эволюции, описаны причины изменения генофонда популяции, открыта роль ДНК как материальной основы наследственности и изменчивости и многое другое.
В синтетической теории эволюции важное место занимает популяционно-генетический подход. Популяционная генетика изучает, как движущие силы эволюции влияют на изменение частот аллелей в популяциях, пространственную структуру популяций, объясняет видообразование.
В 30-х годах XX века в трудах Фишера, Холдейна и Добжанского была показана взаимосвязь между степенью генетической изменчивости популяции и скоростью ее эволюции.
В 1942 г. Хаксли предложил понятие «эволюционный синтез», а в 1949 Симпсон использовал термин «синтетическая теория эволюции».
Эволюция в СТЭ рассматривается как процесс постепенный процесс изменения генофонда популяций (изменяются частоты аллелей разных генов, появляются новые аллели и гены, другие исчезают).
Согласно синтетической теории эволюции элементарной единицей эволюции является популяция.
Под элементарными эволюционными факторами в СТЭ и популяционной генетике понимают процессы, изменяющие набор и частоту аллелей. Выделяют ряд основных эволюционных факторов (несколько по-разному в разных источниках):
Естественный отбор в синтетической теории эволюции рассматривается как самый важный фактор эволюционного процесса. В результате действия естественного отбора в большей степени воспроизводятся наиболее приспособленные к данной среде обитания генотипы. Естественный отбор может быть направлен как против определенных аллелей, так и определенных генотипов (сочетаний аллелей и генов).
Борьба за существование. Дарвин считал ее главным фактором эволюции, а естественный отбор уже следствием борьбы за существование.
Мутационный процесс приводит к возникновению нового генетического материала (новых аллелей генов или даже новых генов). Хотя мутации происходят редко и чаще всего вредны, именно они во многом служат материалом для действия естественного отбора.
Поток генов — это изменение частот аллелей в популяции в результате миграции особей. Кроме того, поток генов приводит к обмену генами между разными популяциями, что уменьшает вероятность их расхождения в процессе видообразования.
Дрейф генов подразумевает случайные изменения частот аллелей и связан с ошибкой выборки, чем и отличается от потока генов. Дрейф генов при «эффекте основателя» заключается в возникновении новой популяции из небольшого количества мигрировавших особей другой популяции. Эти особи не несут весь генофонд исходной популяции, а лишь часть аллелей. В дальнейшем при размножении и увеличении численности генофонд новой популяции будет отличаться от исходной. Другой разновидностью дрейфа генов является «эффект бутылочного горлышка», когда численность популяции резко снижается в результате действия неблагоприятных условий.
Изоляция — возникновение барьеров между популяциями, препятствующих скрещиванию особей и обмену генами. В результате каждая популяция может пойти своим эволюционным путем.
В синтетической теории эволюции показано, как гетерозиготы (несущие обычно вредные рецессивные мутации) служат потенциальным источником процесса эволюции. Рецессивные аллели почти никогда полностью не элиминируются из популяции, а при малом количестве сохраняются в гетерозиготах.
Важное значение в СТЭ уделяют рекомбинации генетического материала. Часто его рассматривают как вторичный эволюционный фактор, возникающий на базе выше перечисленных первичных эволюционных факторов, которые создают изменчивость по отдельным генам, а уже рекомбинацию генов можно рассматривать как вторичный процесс.
Рекомбинация дает большое разнообразие генотипов в популяции даже при малом количестве мутаций. Т. е. при незначительном уровне разнообразия по аллелям наблюдается значительный уровень разнообразия по генотипам (так как генотип состоит из огромного количества генов).
В любом случае мутационная и рекомбинационная изменчивости поставляют материал для естественного отбора.
В результате естественного отбора у популяций и видов появляются адаптации к среде обитания, происходит видообразование (на уровне микроэволюции), возникновение более крупных таксонов (на уровне макроэволюции).

Синтетическая теория эволюции — эволюционная теория(современная), которая опирается на основы генетики и дарвинизма(также на палеонтологию, систематику, молекулярную биологию и др.)
1)элементарной единицей эволюции считается локальная популяция;
2)материалом для эволюции являются мутационная и рекомбинационная изменчивость;
3)естественный отбор рассматривается как главная причина развития адаптаций, видообразования и происхождения надвидовых таксонов;
4)дрейф генов и принцип основателя выступают причинами формирования нейтральных признаков;
5)вид есть система популяций, репродуктивно изолированных от популяций других видов, и каждый вид экологически обособлен;
6)видообразование заключается в возникновении генетических изолирующих механизмов и осуществляется преимущественно в условиях географической изоляции.
- http://studfiles.net/preview/4619345/
- http://studref.com/359608/ekonomika/sinteticheskaya_teoriya_evolyutsii
- http://studopedia.ru/10_180822_tri-vida-rnk.html
- http://bio.wikireading.ru/4356
- http://zen.yandex.ru/media/id/5a77f0bead0f22e83044432c/5aa76bb67ddde8292f6cae48
- http://biology.su/evolution/modern-synthesis
- http://znanija.com/task/963396