ФАСЕТОЧНЫЕ ГЛАЗА — сложные глаза (oculi), основной парный орган зрения ракообразных, насекомых и нек-рых других беспозвоночных, образованный омматидиями, роговичная линза к-рых имеет вид выпуклого 6-гранника — фасетки (франц. facette — грань, отсюда назв.). Ф. г. насекомых неподвижно расположены по бокам головы, иногда (у стрекоз, мух и др.) занимая почти всю её поверхность, у нек-рых ракообразных — на подвижных стеблевидных выростах. Наиб, изучены Ф. г. насекомых, у к-рых они сложены большим числом (до 30 тыс.) омматидиев. Различают 3 морфофункц. типа Ф. г. В аппозиционных Ф. г. (свойственны обычно дневным насекомым) смежные омматидии постоянно изолированы друг от друга непрозрачным пигментом, локализованным в спец. пигментных клетках. В оптикосуперпозиционных Ф. г. изоляция омматидиев переменная, и при недостатке света происходит наложение (суперпозиция) лучей, прошедших сквозь разные фасетки. В таких Ф. г. с «прозрачной зоной», свойственных ночным насекомым и ракообразным, тела зрительных клеток и утолщённый рабдом сдвинуты проксимально. Для нейросуперпозиционных Ф. г. мух характерна суммация нервных сигналов от неск. зрительных клеток, получающих свет из одной и той же точки пространства. Нервная проекция сетчатки на оптич. ганглии мозга и свойства оптики таковы, что Ф. г. обеспечивают анализ внеш. мира с точностью до растра омматидиев, а не отд. зрительных клеток, как у позвоночных. Низкая угловая плотность омматидиев (их оптич. оси расходятся под углом от 1 до 6-8° и больше) препятствует различению мелких деталей, но малая инерционность в сочетании с высокой контрастной чувствительностью сетчатки позволяет нек-рым насекомым воспринимать мелькания с частотой до 250- 300 Гц. Ф. г. обеспечивают мн. беспозвоночным цветовое зрение с восприятием УФ-лучей и анализ направления плоскости линейно поляризованного света, благодаря чему они могут ориентироваться по картине поляризации безоблачного неба. (см. ОММАТИДИИ).
Схема возникновения сетчаточного изображения в аппозиционных (а), оптикосуперпозициониых (6) и нейросуперпозиционных (в) фасеточных глазах: 1 — отдельные омматидии со слитым (а, б) или открытым (в) рабдомом; 2 — аксоны зрительных клеток; 3 — рабдом слитый; 4 — рабдом открытый. Заштрихованы те рабдомы, на к-рые попадают параллельно идущие лучи света (показаны стрелками.).
Глаза фасеточные: чем отличаются от простых?
В процессе эволюции зрения у некоторых животных возникают довольно сложные оптические приборы. К таким, безусловно, можно отнести глаза фасеточные. Они сформировались у насекомых и ракообразных, некоторых членистоногих и беспозвоночных. Чем отличается фасеточный глаз от простого, каковы его основные функции? Об этом поговорим в нашем сегодняшнем материале.
Глаза фасеточные
Это оптическая система, растровая, где отсутствует единая сетчатка. А все рецепторы объединены в небольшие ретинулы (группы), образуя выпуклый слой, не содержащий более никаких нервных окончаний. Таким образом, глаз состоит из множества отдельных единиц – омматидий, объединяющихся в общую систему зрения.
Глаза фасеточные, присущие, к примеру, насекомым, отличаются от бинокулярных (присущих в том числе и человеку) плохим определением мелких деталей. Зато они способны различать колебания света (до 300 Гц), тогда как для человека предельные возможности – 50 Гц. А еще мембрана такого типа глаз имеет трубчатую структуру. Ввиду этого глаза фасеточные не имеют таких особенностей рефракции, как дальнозоркость или близорукость, для них неприменимо понятие аккомодации.
Некоторые особенности строения и зрения
У многих насекомых органы зрения занимают большую часть головы и фактически неподвижны. К примеру, глаза фасеточные у стрекозы состоят из 30 000 частиц, образуя сложную структуру. У бабочек – 17 000 омматидиев, у мухи – 4 тысячи, у пчелы – 5. Наименьшее количество частичек у муравья рабочего – 100 штук.
Бинокулярное или фасеточное?
Первый тип зрения позволяет воспринимать объем предметов, их мелкие детали, оценивать расстояния до объектов и их расположение по отношению друг к другу. Однако бинокулярное зрение человека ограничивается углом в 45 градусов. Если обзор необходим более полный, глазное яблоко осуществляет движение на рефлекторном уровне (либо мы повернем голову вокруг оси). Фасеточные глаза в виде полусфер с омматидиями позволяют видеть окружающую действительность со всех сторон, не поворачивая органов зрения или головы. Причем изображение, которое передает при этом глаз, очень похоже на мозаику: одной структурной единицей глаза воспринимается отдельный элемент, а вместе они отвечают за воссоздание полной картины.
Разновидности
Омматидии имеют анатомические особенности, в результате чего и различаются их оптические свойства (к примеру, у разных насекомых). Ученые определяют три вида фасета:
- Аппозиционные. Такие сложные фасеточные глаза есть у дневных насекомых. Пигмент, не имеющий прозрачных свойств, разделяет фасеты – частички, что находятся рядом. И глазные рецепторы могут воспринимать только свет, совпадающий с осью определенного омматидия.
- Оптикосуперпозиционные. Такие сложные фасеточные глаза имеют некоторые ракообразные, а также ночные и сумеречные насекомые. Пигмент, содержащийся в глазу, попеременно изолирует омматидии, перемещаясь, что повышает чувствительность органов зрения при небольшом освещении.
- Нейросуперпозиционные. Различные омматидии суммируют сигнал, поступающий из одной точки в пространстве.
Кстати, некоторые виды насекомых имеют смешанный тип фасеточных органов зрения, а у многих, кроме рассматриваемых нами, имеются еще и простые глаза. Так, у мухи, к примеру, по бокам головы расположены парные фасеточные органы довольно больших размеров. А на темени есть три простых глаза, выполняющих вспомогательные функции. Такая же организация органов зрения и у пчелы – то есть всего пять глаз!
У некоторых ракообразных фасеточные глаза как бы сидят на подвижных выростах-стебельках.
А у некоторых амфибий и рыб имеется еще и дополнительный (теменной) глаз, который различает свет, но обладает предметным зрением. Сетчатка его состоит только из клеток и рецепторов.
Современные научные разработки
В последнее время глаза фасеточные – предмет изучения и восторга ученых. Ведь такие органы зрения, ввиду своего оригинального строения, дают почву для научных изобретений и изысканий в мире современной оптики. Основные преимущества – широкий обзор пространства, разработка искусственных фасеток, используемых преимущественно в миниатюрных, компактных, секретных системах наблюдения.
Особенности фасеточного зрения насекомых.
В отличие от человека, которому присуще бинокулярное зрение, фасеточные глаза у насекомых неподвижны. У стрекозы глаза занимают почти всю поверхность ее головы и состоят из 25-30 тысяч структурных частиц. Количество омматидиев у разных видов насекомых варьируется: у рабочего муравья их около 100, у пчелы — 5 тысяч, у комнатной мухи — 4 тысячи, у бабочек — до 20 тысяч.
Чем же бинокулярное зрение отличается от фасеточного? Бинокулярное зрение дает возможность воспринимать мир объемным, координировать свои движения при ходьбе, прыжках, ориентироваться на местности, оценивая расстояние от одного объекта до другого и расположение объектов относительно друг друга.
Однако бинокулярное зрение ограничено в пространстве определённым углом зрения (для человека около 50-60°). Если нам нужен больший обзор, мы рефлекторно двигаем глазное яблоко в сторону или поворачиваем голову. Фасеточное зрение насекомых благодаря устройству глаз в виде двух полусфер с большим количеством омматидиев позволяет насекомым видеть предметы и окружающее пространство со всех сторон, не поворачивая головы.
Изображение, воспринимаемое фасеточным типом зрения, выглядит мозаично: каждый элемент воспринимается одной структурной единицей глаза, вместе они воссоздают общую картинку.
Разновидности фасеточных глаз.
Анатомические особенности омматидиев, обеспечивающих фасеточное зрение, и их оптические свойства у разных насекомых отличаются. Различают три типа фасеточных глаз:
• Апозиционные — встречаются у дневных насекомых. Непрозрачный пигмент постоянно разделяет фасетки, находящиеся рядом, поэтому рецепторы глаза воспринимают только тот свет, который совпадает с осью данного омматидия;
• Оптикосуперпозиционные — встречаются у сумеречных и ночных насекомых и ракообразных. Пигмент имеет способность перемещаться и изолировать омматидии попеременно, что повышает чувствительность глаз при слабом освещении;
• Нейросуперпозиционные — зрительные клетки, находящиеся в разных омматидиях, но получающие свет из одной и той же точки пространства, суммируют сигнал.
Особенности фасеточного зрения насекомых, заключающиеся в широком обзоре пространства, дали начало новому направлению в развитии прикладной оптики, направленному на разработку искусственных фасеточных глаз, которые можно использовать в миниатюрных системах видеонаблюдения и контроля.
Цветовосприятие у насекомых сильно различается, однако у большинства, по сравнению с человеком, доступный зрению спектр лучей уменьшен с левой стороны (красный, оранжевый) и увеличен с правой стороны (синий, фиолетовый).
Например, пчелы воспринимают красный, розовый, оранжевый, желтый и зеленый цвета как различные оттенки желтого и не видят между ними большой разницы. Качественно они отличают друг от друга всего 4 цвета. Напротив, бражник различает только две группы цветов: сине-фиолетовую и желто-зеленую. При этом бражники способны полноценно воспринимать эти цвета лишь в сумерках, когда для человеческого глаза все изображение уже сливается в плохо различимые оттенки серого и черного.
Насекомые способны различать формы предметов, но это происходит совсем не так, как у человека. Насекомые, питающиеся нектаром (бабочки, пчелы), игнорируют ‘нерасчлененные’ фигуры (овал, круг, квадрат и др.), их привлекают т.н. ‘расчлененные’ формы: радиальные, напоминающие венчики цветков. Чем сложнее форма и игра теней у предмета, тем лучше он ими воспринимается. Кроме того, пчелы испытывают «тягу» к мелким предметам (например, рисункам на бумаге), обращая на них больше внимания, чем на крупные.
Определенную роль в восприятии формы играет движение объекта. Насекомые охотнее садятся на цветы, которые колышутся на ветру, чем на неподвижные. Личинки стрекоз бросаются за движущейся добычей, а самцы бабочек реагируют на летящих самок и плохо видят сидящих. Вероятно, дело в определенной частоте раздражения омматидиев глаз при движении, мелькании и мерцании.
Насекомые узнают знакомые объекты не только по цвету и форме, но и по расположению предметов, находящихся вокруг них. Например, песчаная оса находит вход в норку, ориентируясь по тем предметам, что располагаются вокруг нее (трава, камни). Если же их убрать или изменить их расположение, это может сбить насекомое с толку.
Эта особенность лучше всего исследована на примере стрекоз, жужелиц и других хищных насекомых. Возможность определять расстояние обусловлена наличием у высших насекомых бинокулярного зрения, то есть двух глаз, поля зрения которых частично пересекаются. Особенности строения глаз определяют, насколько велико расстояние, доступное обзору того или иного насекомого. Например, жуки-скакуны реагируют на добычу и набрасываются на нее, когда находятся от объекта на расстоянии 15 см.
Многие насекомые двигаются так, что у них постоянно сохраняется один и тот же угол падения света на сетчатку. Таким образом, солнечные лучи являются своеобразным компасом, по которому ориентируется насекомое. По тому же принципу ночные бабочки перемещаются в направлении искусственных источников света.
Поделиться в социальных сетях:
Ваше имя:
Разрешены только русские или английские буквы + пробел.
Ваш email:
Содержимое этого поля является приватным и не будет отображаться публично.
Ваш комментарий:
HTML теги и ругательства запрещены. Максимальная длина сообщения 600 символов.
Символьная ASCII CAPTCHA: Обновить
Введите 6 цифр на картинке выше.
Этот вопрос задается для того, чтобы выяснить, являетесь ли Вы человеком или представляете из себя автоматическую спам-рассылку.
Исследование производится в темной комнате, матовая лампа находится на расстоянии 40-50 см впереди больного на уровне его головы. В правой руке врача находится лупа в 13,0 дптр, с помощью которой.
Читать далее.
А ведь врачи-офтальмологи постоянно напоминают: профилактика проблем со зрением – верный способ обезопасить себя от серьезных последствий в будущем. Проверять состояние ваших глаз, как минимум раз в полгода рекомендуется даже тем, кто.
Читать далее.
Расслабляющая гимнастика для глаз: несколько упражнений, которые нужно знать офисным работникам.
Читать далее.
Это метод детального зрительного исследования тканей живого глаза. Метод позволет исследовать передний и задний отделы глазного яблока при различных освещении и величине изображения. Исследование проводят с помощью .
Читать далее.
Глазное яблоко у детей имеет анатомические и физиологические особенности по сравнению с глазами взрослых. В данной статье мы рассмотрим основные характеристики, касающиеся строения глазного яблока у детей. Размер глазного яблока у здорового.
Читать далее.
Информация о графике работы и телефонах всех взрослых и детских больниц города Минска республики Беларусь.
Читать далее.
фасеточные глаза
Сложные глаза (oculi), основной парный орган зрения ракообразных, насекомых и нек-рых других беспозвоночных, образованный омматидиями, роговичная линза к-рых имеет вид выпуклого 6-гранника — фасетки (франц. facette — грань, отсюда назв.). Ф. г. насекомых неподвижно расположены по бокам головы, иногда (у стрекоз, мух и др.) занимая почти всю её поверхность, у нек-рых ракообразных — на подвижных стеблевидных выростах. Наиб, изучены Ф. г. насекомых, у к-рых они сложены большим числом (до 30 тыс.) омматидиев. Различают 3 морфофункц. типа Ф. г.
В аппозиционных Ф. г. (свойственны обычно дневным насекомым) смежные омматидии постоянно изолированы друг от друга непрозрачным пигментом, локализованным в спец. пигментных клетках.
В оптикосуперпозиционных Ф. г. изоляция омматидиев переменная, и при недостатке света происходит наложение (суперпозиция) лучей, прошедших сквозь разные фасетки. В таких Ф. г. с «прозрачной зоной», свойственных ночным насекомым и ракообразным, тела зрительных клеток и утолщённый рабдом сдвинуты проксимально.
Для нейросуперпозиционных Ф. г. мух характерна суммация нервных сигналов от неск. зрительных клеток, получающих свет из одной и той же точки пространства. Нервная проекция сетчатки на оптич. ганглии мозга и свойства оптики таковы, что Ф. г. обеспечивают анализ внеш. мира с точностью до растра омматидиев, а не отд. зрительных клеток, как у позвоночных. Низкая угловая плотность омматидиев (их оптич. оси расходятся под углом от 1 до 6—8° и больше) препятствует различению мелких деталей, но малая инерционность в сочетании с высокой контрастной чувствительностью сетчатки позволяет нек-рым насекомым воспринимать мелькания с частотой до 250— 300 Гц. Ф. г. обеспечивают мн. беспозвоночным цветовое зрение с восприятием УФ-лучей и анализ направления плоскости линейно поляризованного света, благодаря чему они могут ориентироваться по картине поляризации безоблачного неба.
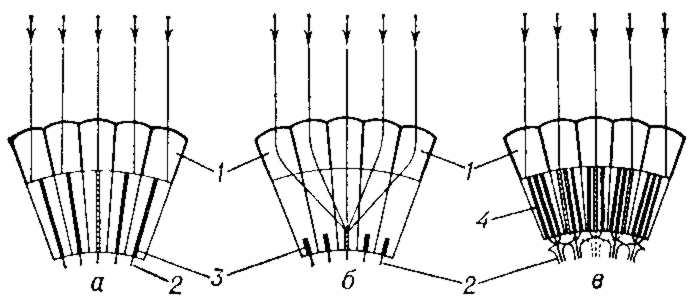
↑ Схема возникновения сетчаточного изображения в аппозиционных (а), оптикосуперпозициониых (6) и нейросуперпозиционных (в) фасеточных глазах: 1 — отдельные омматидии со слитым (а, б) или открытым (в) рабдомом; 2 — аксоны зрительных клеток; 3 — рабдом слитый; 4 — рабдом открытый. Заштрихованы те рабдомы, на к-рые попадают параллельно идущие лучи света (показаны стрелками.).
Значение ФАСЕТОЧНЫЕ ГЛАЗА в Энциклопедии Биология
(сложные глаза), основной парный орган зрения ракообразных, членистоногих и некоторых других беспозвоночных, состоящий из одинаковых простых «глазков» – омматидиев, расположенных в геометрическом порядке. Фасеточные глаза всегда выпуклые. Каждый омматидий даёт прямое изображение части расположенного непосредственно перед ним предмета, а общий его вид складывается, как мозаика, из отдельных частей с участием всех омматидиев. Омматидии членистоногих – мельчайшие светочувствительные органы, состоящие из линзы роговицы, кристаллического конуса, зрительных рецепторных клеток, расположенных, как дольки, в апельсине, и примыкающих к ним пигментных клеток. У различных животных может быть разное количество омматидиев (от нескольких штук до нескольких десятков тыс.). Так, у дафнии 22 омматидия, а у стрекозы ок. 30 тыс. Различают 3 типа фасеточных глаз: аппозиционные, оптико-суперпозиционные и нейросуперпозиционные. В аппозиционных глазах смежные омматидии изолированы друг от друга пигментными клетками. Такие глаза присущи в основном дневным животным, напр. пчёлам, крабам, ракам-богомолам. Оптико-суперпозиционные глаза в условиях сильной освещённости могут функционировать как аппозиционные, но при слабом освещении форма пигментных клеток изменяется, образуется т. н. «прозрачная зона», благодаря чему лучи света из разных омматидиев собираются на один. Подобные глаза присущи ночным ракообразным (креветкам, омарам) и бабочкам. Вершиной эволюции зрения являются нейросуперпозиционные глаза, напр. у мух, в которых разрешающая сила может быть в 100 раз выше, чем в глазах другого типа. Лучи света одного источника падают на зрительные клетки смежных омматидиев, аксоны которых сходятся на одном патроне оптического ганглия. В каждом омматидии 8 зрительных клеток, воспринимающих лучи различных источников света.
Что такое фасеточное зрение в биологии
ФАСЕТОЧНЫЕ ГЛАЗА — сложные глаза (oculi), основной парный орган зрения ракообразных, насекомых и нек рых других беспозвоночных, образованный омматидиями, роговичная линза к рых имеет вид выпуклого 6 гранника фасетки (франц. facette грань, отсюда назв.). Ф. г.… … Биологический энциклопедический словарь
ФАСЕТОЧНЫЕ ГЛАЗА — сложные глаза у некоторых насекомых. Словарь иностранных слов, вошедших в состав русского языка. Чудинов А.Н., 1910. ФАСЕТОЧНЫЕ ГЛАЗА сложные глаза, встречаются у большинства насекомых и состоят из значит. числа простых глазков: у муравьев от 50… … Словарь иностранных слов русского языка
ФАСЕТОЧНЫЕ ГЛАЗА — (от франц. facette грань) (сложные глаза) парный орган зрения насекомых, ракообразных и некоторых др. беспозвоночных; образован многочисленными отдельными глазками омматидиями. Хорошо воспринимают движущиеся объекты, обеспечивают широкое поле… … Большой Энциклопедический словарь
фасеточные глаза — (от франц. facette грань), сложные глаза, парный орган зрения насекомых, ракообразных и некоторых других беспозвоночных; образован многочисленными отдельными глазами омматидиями. Хорошо воспринимают движущиеся объекты, обеспечивают широкое поле … Энциклопедический словарь
Фасеточные глаза — сложные глаза, основной парный орган зрения насекомых, ракообразных и некоторых др. беспозвоночных; образованы особыми структурными единицами – омматидиями (См. Омматидий), роговичная линза которых имеет вид выпуклого шестигранника –… … Большая советская энциклопедия
ФАСЕТОЧНЫЕ ГЛАЗА — (от франц. facette грань) (сложные глаза), парный орган зрения насекомых, ракообразных и нек рых др. беспозвоночных; образован многочисл. отдельными глазами омматидиями. Хорошо воспринимают движущиеся объекты, обеспечивают широкое поле зрения.… … Естествознание. Энциклопедический словарь
Фасеточные глаза — или сложные глаза членистоногих (см. Глаз) получили это название потому, что хитин покровов образует над каждым глазком утолщение, или фасетку (Cornea Linse). Вся совокупность многогранных фасеток представляет поле, напоминающее торцовую мостовую … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
сложные глаза — то же, что фасеточные глаза. * * * СЛОЖНЫЕ ГЛАЗА СЛОЖНЫЕ ГЛАЗА, то же, что фасеточные глаза (см. ФАСЕТОЧНЫЕ ГЛАЗА) … Энциклопедический словарь
СЛОЖНЫЕ ГЛАЗА — то же, что фасеточные глаза … Большой Энциклопедический словарь
сложные глаза — то же, что фасеточные глаза. .(Источник: «Биология. Современная иллюстрированная энциклопедия.» Гл. ред. А. П. Горкин; М.: Росмэн, 2006.) … Биологический энциклопедический словарь
Фасеточные глаза
Фасе́точные глаза́(от франц. facette — грань) — сложные глаза, основной парный орган зрения насекомых, ракообразных и некоторых других беспозвоночных; образованы особыми структурными единицами — омматидиями, роговичная линза которых имеет вид выпуклого шестигранника — фасетки [1] .
Фасеточные глаза насекомых неподвижны, расположены по бокам головы и могут занимать почти всю её поверхность (у стрекоз, мух, пчёл). Фасеточные глаза расположены на капсуле головы в глубоких впячиваниях кутикулы, называемыми глазными капсулами. Кольцо из кутикулы, охватывающее глаз извне, удерживает его на головной капсуле. У ракообразных иногда сидят на подвижных выростах. Наиболее изучены фасеточные глаза взрослых насекомых и их личинок с неполным превращением, у которых они сложены сотнями и даже тысячами омматидиев.
Глаза различных видов насекомых состоят из различного числа омматидиев: у рабочего муравья — около 100, у комнатной мухи — около 4000, у рабочей пчелы — 5000, у бабочек — до 17 000, у стрекоз — до 30 000.
Содержание
Типы фасеточных глаз [ править ]
В зависимости от анатомических особенностей омматидиев и их оптических свойств различают 3 типа фасеточных глаз: апозиционные (фотопические), оптикосуперпозиционные и нейросуперпозиционные (называемые в совокупности скотопическими). У некоторых насекомых (богомолы, подёнки) одна часть глаза может быть построена по аппозиционному типу, а другая — по суперпозиционному.
В фасеточных глазах всех типов собственно светочувствительным элементом служат рабдомеры зрительных клеток, содержащие фотопигмент (обычно подобный родопсину). Поглощение фотопигментом квантов света — первое звено в цепи процессов, в результате которых зрительная клетка генерирует нервный сигнал.
Апозиционные (фотопические) фасеточные глаза [ править ]
В апозиционных фасеточных глазах, свойственных обычно дневным насекомым, смежные омматидии постоянно изолированы друг от друга непрозрачным пигментом и рецепторы воспринимают только свет, направление которого совпадает с осью данного омматидия.
Оптикосуперпозиционные фасеточные глаза [ править ]
В оптикосуперпозиционных фасеточных глазах, характерных для ночных и сумеречных насекомых и многих ракообразных, изоляция омматидиев переменная (вследствие способности пигмента перемещаться), и при недостатке света происходит наложение (суперпозиция) падающих под косым углом лучей, прошедших не сквозь одну, а сквозь несколько фасеток. Таким образом, при слабом освещении увеличивается чувствительность глаза.
Нейросуперпозиционные фасеточные глаза [ править ]
Для нейросуперпозиционных фасеточных глаз характерна суммация сигналов от зрительных клеток, находящихся в разных омматидиях, но получающих свет из одной и той же точки пространства.
Разрешающая способность и цветовое восприятие [ править ]
Нервная проекция сетчатки на оптические ганглии мозга и, отчасти, особенности оптики фасеточных глаз таковы, что они обеспечивают анализ внешнего мира с точностью до линзового растра омматидиев, а не отдельных зрительных клеток. Низкая угловая плотность омматидиев (их оптические оси расходятся под углами 1—6°) препятствует получению изображения объекта на относительно близком расстоянии (принцип работы телеобъентива), но дают возможность различать мелкие детали на достаточном расстоянии для оценки и запоминания обстановки. Например, пчёлы, стрекозы и др., обладая строением фасеточной зретильной системы (растровое) в виде двух «полусфер», обеспечивают стереоскопическое зрение в допустом поле зрения и практически общее зрение в поле сферы! без повора головы. (Данная система зрения представляет большой интерес в науке, в военном деле, медицине и др. областях). Малая инерционность в сочетании с высокой контрастной чувствительностью (1—5 %) фасеточных глаз позволяет некоторым насекомым различать мелькания (мигания) света с частотой вплоть до 250—300 Гц (для человека предельная частота около 50 Гц). Фасеточные глаза обеспечивают многим беспозвоночным цветовое зрение с восприятием ультрафиолетовых лучей, а также анализ направления плоскости линейно-поляризованного света [2] .
Использование строения фасеточных глаз в науке [ править ]
Исследователи пытались копировать систему фасеточного зрения, когда направленные в разные стороны несколько сегментов глаза с небольшим разрешением, но с полем зрения практически сферическом, образуют в мозгу насекомого цельную картинку обстановки вркруг себя без поворота головы. При этом уникалность такой биологической зрительной системы состоит в том, что она обеспечивает припосабливаемость обитателей к внешней среде сосуществования при самом рациональном механизме построения. (Вообще, чем больше и ближе мы приближаемся к природе, познаём её, тем точнее нам удаётся создавать необходимые для себя открытия. При этом нельзя упрощать всё, что создано природой). До сих пор подобные системы были слишком велики из-за большого числа линз. В иных случаях для расширения угла обзора используются громоздкие линзы типа «рыбий глаз», специальный механизм для поворота объектива или поворотные зеркала, как в оптических устройствах для эндоскопии.
Инженерам из британской обороны удалось создать компактное решение с фасеточным зрением. В новой камере используется всего 9 линз, соединенных со стационарным сенсором изображения с помощью миллионов оптических волокон. Каждая из линз направляет полученное изображение на свой участок фотосенсора. Для склеивания фрагментов изображения в единую картину используется специальная программная система.
Новая камера, изначально предназначенная для систем наведения в боевых ракетах, имеет размер не больше кусочка растворимого сахара. По мнению разработчиков, подобные камеры могут найти широкое применение в микрохирургии, так как нынешние камеры с поворотными зеркалами подвержены частым отказaм из-за большого числа движущихся частей. Кроме того, новая фасеточная система компании BAE Systems может существенно расширить возможности обычных камер видеонаблюдения.
Эксперты по зрению насекомых отмечают, что фасеточная система, используемая в новых камерах, потенциально повышает устойчивость решения к отказам и повреждениям (даже если одна линза будет повреждена, остальные могут взять на себя часть ее функций) [3] [4] [5] .
Фасеточные глаза, фасеточные глаза — сложные глаза, основной парный орган зрения насекомых, ракообразных и некоторых др. беспозвоночных; образованы особыми структурными единицами — омматидиями, afctnjxyst ukfpf энциклопедия биология экология животные раст
Тип членистоногие (arthropodа)
Среди всех групп беспозвоночных животных тип членистоногих выделяется наибольшим разнообразием приспособлений к самым различным условиям существования, изумительным богатством форм и огромным числом видов. Число видов членистоногих приближается к 3 миллионам и намного превышает число видов всех остальных типов животных и растений, вместе взятых, включая микроорганизмы. Особенно обильны видами насекомые, на долю которых падает более 90% известных видов членистоногих.
Именно тонкими приспособлениями к использованию всевозможных местообитаний, всяких источников пищи, часто недоступных для других животных, объясняется многообразие и обилие видов членистоногих.
Трудно найти такие места, где не было бы членистоногих. Они живут во всех морях и океанах — и в толще воды, и на дне, и в грунте на разных глубинах; они обитают и во всевозможных пресных водоемах — не только в реках и озерах, но и в мелких пересыхающих лужах, в подземных водах и пещерах, в скоплениях воды в дуплах деревьев. Некоторые виды приспособились к жизни в очень соленых водоемах, другие — в горячих источниках. Кажется, все разнообразие водной среды на поверхности нашей планеты освоено членистоногими.
Большая армия представителей этого типа приспособилась и к жизни вне воды — на суше, где членистоногие особенно многообразны. Весь слой почвы, пронизанный корнями растений, обильно заселен и членистоногими — во многих лесных и луговых почвах миллион особей клещей и ногохвосток на 1 м2 не редкость. Огромное количество членистоногих обитает на поверхности различных частей растений и внутри растительных тканей.
Во всех климатических зонах можно встретить членистоногих. Огромно их количество и велико многообразие в тропиках, но есть они и в высоких широтах Арктики и на побережье Антарктиды, где недавно были обнаружены новые виды клещей и ногохвосток. Членистоногих можно встретить и в самых сухих пустынях. Образно можно сказать, что членистоногие — это тип животных, который по полноте овладения поверхностью земного шара не имеет равных.
По многим признакам строения членистоногие близки к кольчатым червям, особенно к полихетам. Замечательный зоолог начала прошлого века Жорж Кювье, предложивший деление животных на типы, объединил кольчатых червей и членистоногих в единый тип членистых животных (Articulata). Знаменательно, что и в наши дни некоторые авторитетные зоологи, например В. Н. Беклемишев, ратуют за восстановление типа членистых. Это показывает, что членистоногие во всяком случае очень близки к многощетинковым кольчатым червям. От близких к ним форм когда-то давно, еще в те геологические эпохи, в отложениях которых не сохранилось ископаемых остатков, отделились предки современных членистоногих.
Членистоногие, как и кольчатые черви, — это двустороннесимметричные сегментированные животные. Сходство членистоногих с кольчецами проявляется не только в наружной сегментации. Центральная нервная система у более примитивных членистоногих очень сходна с лестничной нервной системой полихет.
Специфические признаки членистоногих связаны с тем, что у них тело покрыто очень плотной и прочной кутикулой, которая нередко образует очень твердый панцирь. Кутикула у членистоногих выделяется наружным слоем клеток, так называемой гиподермой, имеет очень сложное строение и сложный химический состав. Раньше считали, что у членистоногих кутикула состоит только из хитина. Теперь выяснилось, что, кроме хитина, в состав ее входят многие другие соединения, в основном задубленные белки, и что у большинства членистоногих на долю хитина падает не больше трети веществ, из которых состоит кутикула. Хитин — это сложный полисахарид, в состав молекулы которого входят атомы азота. Вероятно, это не определенное соединение, а группа близких соединений, формулы которых точно не установлены. Хитин эластичен, прочен и устойчив к химическим воздействиям. Нередко в кутикуле содержится много углекислого кальция.
Прочность и твердость кутикулы членистоногих настолько велики, что в тех местах, где она достигает большой толщины, покровы становятся негибкими. Подвижность сегментов тела друг относительно друга, обеспечивающая движение, у членистоногих достигается тем, что между твердыми участками (склеритами) сохраняются перепончатые участки с очень тонкой кутикулой; такие перепонки на сгибах образуют складки, расправляющиеся при растягивании тела. Далеко не все сегменты у членистоногих подвижны друг относительно друга, очень часто наблюдается полное слияние соседних сегментов, особенно в передних отделах тела.
В теле членистоногих обычно можно различить 3 отдела — голову, на которой находятся основные органы чувств, служащие для ориентировки в пространстве (глаза, усики и др.)» и органы захватывания пищи. Сегменты головы у членистоногих слитные. Далее следует грудь — тот отдел тела, на котором располагаются основные двигательные придатки — ноги, а у насекомых и крылья. Задний отдел тела называется брюшком. У некоторых членистоногих (например, у многоножек, а из ископаемых — у трилобитов) все туловищные сегменты более или менее однородны, и тогда в теле животного различают только 2 отдела — голову и туловище. У многих членистоногих (у паукообразных, десятиногих раков) сливаются головной и грудной отделы, образуя головогрудь. А у клещей вообще утрачиваются границы между отделами тела, тело слитное.
Передвигаются членистоногие с помощью конечностей.
Поскольку конечности у них, как и все тело, покрыты кутикулой, их движение возможно тоже только при разделении ног на членики. По строению конечностей, на которые опираются членистоногие при движении по твердой поверхности или с помощью которых они плавают, весь тип и получил свое название.
Некоторое количество пар конечностей на голове у всех членистоногих превращается в ротовые органы — челюсти, использующиеся хотя бы частично для захватывания или размельчения пищи. Это тоже признак, характерный для типа членистоногих. Ротовые части бывают настолько видоизменены, что их трудно даже признать конечностями, например, хоботок комара или клопа. Число пар ног, которые служат для передвижения, неодинаково у разных членистоногих. Для насекомых характерно развитие 3 пар двигательных конечностей, для паукообразных — 4 пар, а у некоторых многоножек число ног превышает сотню.
Нередко туловищные конечности служат не только как ноги, но и как жабры, присоски и другие органы.
У членистоногих двигательная мускулатура состоит из отдельных мышц, концы которых присоединяются к разным утолщенным участкам покровов, отделенным друг от друга мягкой перепонкой. Движение членистоногих обеспечивается не изгибами тела при сокращении всей мускулатуры туловища, как у червей, а сокращениями обычно немногих мышц, связанных с конечностями или другими двигательными придатками (крылья насекомых, вилочка ногохвосток и др.)- Мышцы членистоногих обладают исключительно высокой способностью к сокращению. Все они поперечнополосатые, причем обнаруживаемая под микроскопом поперечная исчерченность мышечных волоконец у членистоногих выражена больше, чем у позвоночных, — мышцы членистоногих сильнее наших, если, конечно, учитывать размеры. Так как панцирь членистоногих служит и местом прикрепления двигательных мышц, он является и скелетом, называемым «наружным» в отличие от привычного нам внутреннего скелета позвоночных.
Панцирь членистоногих лишь в результате долгой эволюции стал скелетом. А сначала утолщение и упрочнение покровов у обитавших в морях далеких червеобразных предков членистоногих служило, надо полагать, приспособлением к защите внутренних органов от повреждений. Утолщение и уплотнение кутикулы делает ее менее проницаемой. Поэтому понятно, что только очень мелкие членистоногие, имеющие тонкие покровы, живущие в воде или в очень влажных местах, могут дышать всей поверхностью тела.
У всех более крупных членистоногих, имеющих и более плотные покровы, обязательно развиваются специальные дыхательные органы, поверхность которых покрыта такой тонкой кутикулой, что через нее легко может поступать кислород.
У живущих в воде членистоногих это жабры. Для дыхания в воде важно, чтобы поверхность жабр была возможно большей. Поэтому жабры всегда представляют собой тонкие листовидные выросты или пучки пальцевидных отростков. Обычно жабрами у членистоногих становятся конечности или части конечностей. В воде благодаря большой ее плотности даже тоненькие жаберные листочки не слипаются друг с другом. Поэтому на небольшом участке тела (в остальных местах защищенного плотными покровами) может разместиться много листовидных жабр.
Для доставки кислорода от жабр ко всем удаленным от них органам служит кровеносная система и полостная жидкость. У членистоногих кровеносная система представлена расположенным в спинной части тела пульсирующим, обычно продолговатым сосудом — сердцем. Кровь поступает в сердце через закрывающиеся клапанами боковые отверстия, а выталкивается из него в полость тела или непосредственно, или через немногие крупные сосуды — система кровообращения незамкнутая. У крупных морских членистоногих, имеющих толстый панцирь, — у омаров, у мечехвостов — в крови есть дыхательный пигмент гемоцианин, сходный по химическому составу с нашим гемоглобином, но придающий крови не красный, а синеватый цвет: в состав гемоцианина входит не железо, как в гемоглобине, а медь. Гемоцианин легко присоединяет кислород при прохождении крови через жабры и отдает его внутренним органам. Так как у членистоногих кровеносная система незамкнутая, правильнее называть их «кровь» «гемолимфой», но и термином «кровь» зоологи широко пользуются.
На открытой поверхности суши ни кожное дыхание, ни дыхание с помощью жабр невозможно легко проницаемые участки покровов пропускают воду и быстро подсыхают, теряя проницаемость. Поэтому водные организмы, извлеченные на сушу, погибают от потери воды или от недостатка поступающего кислорода. На суше только скрытоживущие мелкие членистоногие, обитающие в таких местах, где воздух всегда насыщен водяным паром, например в почве, могут дышать всей поверхностью тела (мелкие клещи, ногохвостки, симфилы). Жизнь на суше вне влажных укрытий требует развития непроницаемых для испарения покровов, а такие покровы не пропускают и кислород.
У наземных членистоногих развиваются особые дыхательные органы — либо «легкие», представляющие глубокие мешковидные впячивания со складчатыми стенками, открывающиеся наружу лишь небольшими отверстиями (у пауков, у скорпионов), либо (у большинства наземных членистоногих) трахеи. Трахеи представляют собой тонкие, обычно обильно ветвящиеся трубочки, открывающиеся наружу маленьким входным отверстием («дыхальцем») на поверхности тела. Ветви этих трубочек омываются полостной жидкостью, а самые мелкие ответвления доходят до отдельных клеток. Из тонких трубочек трахей вода через дыхальца испаряется очень медленно, и потери ее незначительны, а кислород по ним проникает быстро и легко.
Уже далекие предки разных групп членистоногих приспособились к жизни в разных средах. Можно выделить группу водных членистоногих, к которым относятся ракообразные, выделяемую в подтип жабродышащих. Правда, среди современных ракообразных есть и такие, которые живут на суше (мокрицы), но по всем признакам они очень близки к своим водным родичам. Другой подтип — трахейнодышащие — представлен типично наземными членистоногими — различными группами многоножек и насекомых. Они прекрасно приспособлены к жизни на суше. В их покровах содержатся воско- и жироподобные вещества, защищающие организм от высыхания, а все системы органов, связанных с расходованием воды, — выделительная и дыхательная — действуют так, что влага расходуется минимально. Среди насекомых есть отдельные виды, проводящие всю жизнь или часть ее в воде, но строение таких водных насекомых явно показывает их происхождение от обитавших на суше предков.
За длившуюся сотни миллионов лет историю эти группы обособились в связи с приспособлениями к жизни в двух разных средах — в воде и на суше.
Третий подтип членистоногих — хелицеровые — очень существенно отличается и от ракообразных, и от трахейнодышащих. Среди хелицеровых есть и водные представители — мечехвосты, обитающие в морях, и наземные — паукообразные.
В ископаемом состоянии сохранилось много остатков хелицеровых. Их изучение, дополняющее изучение современных представителей, показывает, как шло разделение классов в результате приспособления к жизни в разных средах—в воде и на суше. Изучение всех групп членистоногих убеждает в том, что переход их к жизни на суше был связан со способностью к выработке предохраняющих от испарения влаги покровов, к сохранению в теле воды, без которой невозможна жизнь.
У членистоногих, хорошо приспособившихся к жизни на суше, панцирь бывает нередко тоньше, чем у обитающих в воде, но зато на его поверхности отлагается очень тонкий слой воскоподобных и жироподобных веществ, не пропускающих пары воды. Этот слой защищает организм от высыхания и препятствует испарению. Поэтому у всех членистоногих, живущих открыто на поверхности суши, есть специальные органы дыхания. Те членистоногие, у которых специальных органов дыхания нет, которые дышат всей поверхностью кожи, вынуждены обитать во влажных укрытиях — в почве, в лесной подстилке ит. д., только иногда по ночам и после сильных дождей показываясь на поверхности.
У членистоногих, наилучшим образом приспособившихся к жизни на суше, чаще всего развивается трахейная система. Не только дыхательная, но и другие системы органов меняются у членистоногих с переходом к жизни на суше.
Выведение растворенных вредных продуктов разрушения белков у водных членистоногих происходит непосредственно во внешнюю среду через отверстия выделительных органов, открывающиеся у основания конечностей головного или грудного отдела. А у всех членистоногих, наилучшим образом приспособившихся к жизни на суше, — у пауков, многоножек, насекомых — выделительные трубочки («мальпигиевы сосуды») открываются в начале задней кишки. По мере прохождения по ней непереваренных остатков пищи и продуктов выделения и из тех и из других стенки задней кишки всасывают воду, столь труднодоступную для жителей суши, а экскременты и продукты обмена выводятся из кишечника в обезвоженном состоянии.
Приспособившиеся к жизни на суше членистоногие выработали способность запасать много жира в особых образованиях— в жировом теле. Главная роль жирового тела состоит в том, чтобы в критические периоды, когда организм не может получить воду извне, снабжать его. водой!
При окислении жиров образуется много воды (так называемая «метаболическая вода»), которая и идет на покрытие расхода влаги животным. В этом отношении насекомые сходны с верблюдами, у которых при длительных переходах через пустыню используется вода, образующаяся при окислении запасов жира в горбу. У водных групп членистоногих (ракообразных, мечехвостов) жирового тела не бывает.
Наружный панцирь членистоногих настолько плотен, что в затвердевшем состоянии не растягивается. Поэтому рост членистоногих сопровождается линьками. У растущего животного старый панцирь становится тесным, лопается, обычно сначала на спине, и сбрасывается. И только в короткий период, пока новый панцирь, образовавшийся под ним, не затвердеет, животное может увеличиваться в размерах. Во время сбрасывания старых покровов, которые сходят не только со всей наружной поверхности тела, но и со всех конечностей, в том числе и с ротовых органов, а также с передней и задней кишки и со всех дыхательных органов, членистоногие бывают вялыми, малоподвижными, не питаются и стараются спрятаться в каких-либо укрытиях.
Рост у членистоногих — не непрерывный процесс, а ступенчатый, размеры тела увеличиваются скачкообразно — сразу после линьки. У многих членистоногих во время линьки происходит и большее или меньшее изменение строения тела. Часто взрослое животное приспособлено к совсем иному образу жизни, чем молодое, и потому особи разных возрастов (возрастом у членистоногих называется период между линьками) бывают даже совсем непохожи друг на друга. В таком случае развитие сопровождается метаморфозом. Превращение гусеницы в куколку, а куколки в бабочку может служить общеизвестным примером.
Образ жизни разных членистоногих неодинаков. Многие из них отличаются очень сложным поведением, что всегда связано с высоким развитием органов чувств и центральной нервной системы.
У большинства членистоногих хорошо развиты глаза. Глаза у членистоногих бывают простые, имеющие одну линзу, и сложные, или фасеточные. У высших представителей всех подтипов глаза сложные, фасеточные, состоящие из множества расположенных плотно рядом друг с другом маленьких глазков. Иногда число глазков (так называемых омматидиев) в фасеточном глазу измеряется несколькими тысячами (рис. 185). Фасеточные глаза характерны для высших ракообразных, мечехвостов и насекомых. Такие глаза были и у живших в палеозойскую эру трилобитов — членистоногих, полностью вымерших в конце этой эры. Каждый омматидий воспринимает только одну точку находящегося перед ним предмета (рис. 186). В результате сложный глаз дает изображение, состоящее из большого числа отдельных точек, т. е. мозаичное (как тоновое клише в книге) и прямое (рис. 7, см. т. 3, табл. 18).
Вспомним, что изображение, получаемое в нашем глазу, перевернутое и цельное!
 Глаза членистоногих, оказывается, могут воспринимать не только те лучи, которые видимы для нашего глаза, но и ультрафиолетовую часть спектра. Например, многие цветы, которые нам кажутся равномерно белыми, для насекомых оказываются пестрыми; у таких цветов есть не видимый нами ультрафиолетовый рисунок (см. т. 3, табл. 19).
Глаза членистоногих, оказывается, могут воспринимать не только те лучи, которые видимы для нашего глаза, но и ультрафиолетовую часть спектра. Например, многие цветы, которые нам кажутся равномерно белыми, для насекомых оказываются пестрыми; у таких цветов есть не видимый нами ультрафиолетовый рисунок (см. т. 3, табл. 19).

И наземные и водные членистоногие способны различать обычный свет и так называемый поляризованный (те световые лучи, электромагнитные колебания которых совершаются в одной плоскости). Поскольку для рассеянного света неба характерна определенная степень поляризации лучей в меридиональной плоскости, членистоногие по небу, как по компасу, могут ориентироваться и в ясные и в пасмурные дни. Таким образом, членистоногие различают не только частоту, но и плоскость колебаний световых лучей.

Мы не ощущаем магнитные волны. А например, самки термитов в гнезде лежат так, что ось их тела направлена по магнитному меридиану, а если поместить рядом сильный магнит, меняют свое положение. Как воспринимают членистоногие магнитные волны, пока не выяснено. Отмечено, что членистоногие могут реагировать и на короткие радиоволны.
Хорошо развит и слух у членистоногих. Звуковые волны определенной длины (чаще короткие) они воспринимают с помощью тонких слуховых волосков на поверхности тела, особенно на усиках, с помощью специальных слуховых органов, напоминающих по строению струну, натянутую над резонаторной ямкой (как струна на балалайке), и других приспособлений. Часто наземные членистоногие издают ультразвуки, которых мы не слышим, с помощью различных «стридуляционных» органов, по принципу действия напоминающих струну и смычок.
Особенно замечательна способность членистоногих воспринимать запахи — ничтожные концентрации многих веществ, растворенных в воде или взвешенных в воздухе. Никакие наши химические приборы не дают возможности уловить такие слабые концентрации вещества, а насекомые, например, могут по ним ориентироваться. Бабочки совки могут прилетать за километр на тарелку с бродящей патокой, а самцы шелкопрядов летят за километры на не ощущаемый нами и вблизи запах коробки, в которой сидела самка. Не менее тонко восприятие и запахов, и содержания солей водными членистоногими. Многие наземные членистоногие (мокрицы, личинки насекомых) способны улавливать разницу в относительной влажности воздуха порядка 1—2%.
У водных членистоногих прекрасно развиты органы равновесия.
Наиболее универсальное чувство, присущее всем членистоногим, —осязание. Осязательную функцию выполняют многочисленные волоски на поверхности тела членистоногих. Для многих членистоногих, особенно для донных, почвенных и живущих в тканях растений и животных, характерно стремление к наибольшей поверхности контакта с твердыми предметами — «тигмотаксис».
Многообразная «информация», получаемая из внешнего мира органами чувств членистоногих, требует «обработки информации» центральной нервной системой. Головной мозг — парные скопления нервных клеток над пищеводом — у высших активных членистоногих очень хорошо развит, в нем различается несколько отделов, связанных с основными органами ориентировки в пространстве, расположенными в головном отделе. Чем сложнее поведение членистоногого, чем многообразнее те условия, с которыми животное сталкивается, и чем сложнее ответы организма на воздействия среды, тем более сложно строение головного мозга. Головной мозг у членистоногих связан окологлоточным нервным кольцом с брюшной нервной цепочкой. Исходным у членистоногих, по-видимому, является соответствие каждому сегменту тела пары ганглиев брюшной нервной цепочки. Но при слиянии отдельных сегментов и даже отделов тела друг с другом происходит уменьшение числа нервных узлов, частично сливающихся друг с другом, частично исчезающих. Степень расчленения или слитности брюшной цепочки связана со степенью расчленения или слитности тела, и концентрация узлов брюшной нервной цепочки совсем не может служить показателем сложности поведения.
Поведение членистоногих, особенно в период размножения (спаривание, забота о потомстве), бывает очень сложным, в чем можно убедиться из знакомства с приводимыми ниже описаниями образа жизни разных представителей этого типа. Очень часто поведение определяется прямой реакцией на раздражение, например приближение к источнику света («положительный фототаксис») или уход от света («отрицательный фототаксис»). Еще более распространены реакции на раздражение путем усиления или ослабления подвижности («кинезисы»). Например, многие насекомые активнее двигаются при более сильном или слабом освещении и малоподвижны при среднем. Таким образом, после ненаправленного быстрого движения они, наконец, попадают в благоприятные для себя условия освещения и перестают двигаться.
Кроме непосредственных стандартных ответов на раздражения (безусловных рефлексов), для членистоногих очень характерны и сложные цепи наследственно закрепленных рефлекторных действий (инстинкты). Инстинкты представляют такое сложное поведение, какое в нормальных для данного вида условиях является целесообразным, но оно остается неизменным и при изменении условий и поэтому может становиться нецелесообразным.
Например, роющие осы вырывают норку, приносят в нее парализованное насекомое, которым должна питаться личинка, откладывают на добычу яйцо и заделывают норку. Все действия осы «целесообразны». Но если в процессе деятельности осы, когда она приступает к заделыванию норки, вынуть и добычу и яйцо, оса завершает свою уже ненужную работу по замуровыванию норки.
Наряду со сложными формами инстинктивного поведения у членистоногих (высшие ракообразные, пауки, высшие насекомые) прослеживаются и индивидуальные навыки; у них можно выработать и условные рефлексы. Мало того, например, пчелы могут передавать друг другу приобретенные навыки, обмениваться информацией. Определенными телодвижениями рабочая пчела сообщает другим пчелам в улье, в каком направлении надо лететь за взятком.
Членистоногим, несомненно, свойственны многие проявления высшей нервной деятельности, которую еще недавно считали характерной только для позвоночных.
Типы оплодотворения у членистоногих разнообразны.
У водных членистоногих, например, мечехвостов, бывает настоящее наружное оплодотворение, когда самка откладывает яйца в воду, а самец туда же выводит семенную жидкость. Проникновение сперматозоида в яйцо происходит при наружном оплодотворении во внешней среде — в воде.
Часто при наружном оплодотворении у водных членистоногих самец прикрепляет «пакеты» с семенной жидкостью (сперматофоры) к выводным половым отверстиям самок — откладываемые яйца оплодотворяются во внешней среде, но вероятность оплодотворения каждого яйца в таком случае увеличивается.
С переходом к жизни на суше низшие членистоногие, обитающие во влажной среде, — в почве, в гнилой древесине и т. п. (многие клещи, скорпионы, ложноскорпионы, многоножки, ногохвостки и др.), выработали новый способ оплодотворения. Самцы откладывают сперматофоры во внешнюю среду, как при наружном оплодотворении (на землю, на остатки растений), а самки подбирают их своими половыми отверстиями, и, таким образом, проникновение сперматозоидов в яйцо осуществляется внутри организма самки, как при внутреннем оплодотворении. Такое оплодотворение называется наружно-внутренним.
У обитающих на поверхности суши в открытой атмосфере членистоногих оплодотворение внутреннее: самцы вводят семенную жидкость непосредственно в половые отверстия самок. Проникновение сперматозоида в яйцо происходит в защищенных от высыхания условиях внутри материнского организма.
В отдельных группах водных членистоногих, например у усоногих раков, также выработалась способность к внутреннему оплодотворению, повышающая плодовитость.
У большинства членистоногих наблюдается откладка яиц, живорождение наблюдается редко, например у некоторых двукрылых насекомых. Чаще наблюдается яицеживорождение: самка рождает живых детенышей, однако развитие детенышей происходит за счет питательных запасов яйца, но не во внешней среде, а в яйцеводах самок (например, у тлей).
Как упоминалось, развитие многих членистоногих происходит со сложным превращением. В тех случаях, когда взрослая особь и форма, вышедшая из яйца, резко различаются по строению, молодые особи называются личинками.
У малоподвижных или неподвижных морских ракообразных расселяются подвижные личинки. У очень подвижных во взрослом состоянии крылатых насекомых личинки—в основном питающаяся стадия.
В тех случаях, когда яйца богаты желтком, из них выходят молодые членистоногие, уже похожие на взрослых. Такое развитие характерно для многих пресноводных ракообразных, паукообразных и некоторых насекомых.
Членистоногие — древние животные.
Самые достоверные данные, на основании которых можно судить о происхождении тех или иных групп животных, дают исследования ископаемых окаменевших остатков организмов или их отпечатков в слежавшихся и окаменевших наслоениях ила, песка или глины, под наносами которых трупы животных были погребены. Но не от всех прошедших эпох, когда уже существовала на Земле жизнь, сохранились ископаемые остатки организмов. Отложения самых древних периодов развития жизни на Земле подвергались в течение последующих эпох таким сильным изменениям, что точно установить, какие организмы обитали на Земле около миллиарда лет тому назад, невозможно, сколько-нибудь уцелевших остатков животных из отложений более древних, чем палеозойские, не сохранилось. А уже в самых глубоких («нижних») осадочных породах кембрийского периода палеозойской эры (отлагавшихся более полумиллиарда лет назад) встречаются трилобиты (см. т. 3, рис. 3)— животные с явно выраженными признаками членистоногих, имевшие развитой наружный скелет, расчлененные конечности, сложные глаза и другие свойственные представителям типа Arthropoda признаки организации. Трилобиты— класс членистоногих, полностью вымерший в конце палеозойской эры (длившейся 300—350 миллионов лет).

Из кембрийских отложений известны и ракообразные, некоторые из которых относятся к современным отрядам, с силурийского периода известны родичи мечехвостов (см. т. 3, табл. 1) и даже скорпионов (см. т. 3, табл. 3, 1, 2), в девонских отложениях были найдены остатки ногохвосток, а недавно московский палеонтолог Б. Б. Родендорф установил, что в девоне жили и крылатые насекомые.

Все эти данные свидетельствуют о том, что тип членистоногих отделился от каких-то животных, похожих на кольчецов, очень давно. Поэтому точно установить родственные связи различных членистоногих очень трудно. Несомненно, что одной из первых оформившихся групп членистоногих были трилобиты. В палеозое трилобиты достигли расцвета, они буквально кишели в прибрежной зоне морей и океанов 500 миллионов лет назад. Трилобиты сохранились в ископаемом состоянии настолько хорошо, что по некоторым остаткам удалось не только исследовать их внутреннее строение, но на шлифах изучить даже микроскопическую структуру тканей этих животных, а также проследить их развитие. А применяя некоторые методы современной химии, удалось даже выяснить, что в их крови был гемоцианин, как у современных высших ракообразных.
Трилобиты, обычно выделяемые в отдельный класс членистоногих, заслуживают выделения в самостоятельный подтип Trilobita.
Из современных групп членистоногих наиболее близки к трилобитам, по-видимому, мечехвосты (см. т. 3, табл. 1), личинки которых несколько сходны с трилобитами, а зародыши—с личинками трилобитов («протаспис»—см. т. 3, рис. 2).

Мечехвосты и паукообразные имеют ряд общих черт строения, позволяющих их объединить в единый подтип хелицеровых. (Chelicerata).
По характеру органов дыхания и некоторым другим признакам некоторые общие с трилобитами черты имеют и ракообразные (класс Crustacea). Однако непосредственных родственных связей между трилобитами и ракообразными установить нельзя. Ракообразные представляют особый подтип жабродышащих (Branchiata) в типе членистоногих. А так как среди современных ракообразных многие имеют более примитивное строение, чем хелицеровые, принято рассмотрение типа членистоногих начинать с них.
Последний подтип — трахейнодышащие (Tracheata)—объединяет различных многоножек и насекомых, т. е. типично наземных членистоногих.
В настоящее время не вызывает сомнений происхождение насекомых от каких то древних членистоногих, близких к многоножкам, а многоножек — от примитивных червеобразных членистоногих.
Вероятно, водные хелицеровые произошли от общих предков с трилобитами — это была другая ветвь эволюционного развития членистоногих. А приспособившиеся к жизни в водной среде ракообразные, сохранившие много черт древних членистоногих, представляют третью основную ветвь развития членистоногих (рис. 188).

Знакомство с типом и с его многообразными представителями удобнее всего начинать с подтипа жабродышащих, с класса ракообразных.

Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева.1970.
Фасеточные глаза , сложные глаза, основной парный орган зрения насекомых , ракообразных и некоторых др. беспозвоночных ; образованы особыми структурными единицами — омматидиями, роговичная линза которых имеет вид выпуклого шестигранника — фасетки (франц. facette — грань; отсюда название). Фасеточные глаза насекомых неподвижны, расположены по бокам головы и могут занимать почти всю се поверхность (у стрекоз , мух, пчел). У ракообразных иногда сидят на подвижных выростах. Наиболее изучены Ф. г. взрослых насекомых и их с неполным превращением, у которых они сложены сотнями и даже тысячами омматидиев. В зависимости от анатомических особенностей омматидиев и их оптических свойств различают 3 фасеточных глаз. В аппозиционных Ф. г., свойственных обычно дневным насекомым, смежные омматидии постоянно изолированы друг от друга непрозрачным пигментом и рецепторы воспринимают только свет, направление которого совпадает с осью данного омматидия. В оптикосуперпозиционных Ф. г., характерных для ночных и сумеречных насекомых и многих ракообразных, изоляция омматидиев переменная (вследствие способности пигмента перемещаться), и при недостатке света происходит наложение (суперпозиция) падающих под косым углом лучей, прошедших не сквозь одну, а сквозь несколько фасеток. Таким образом, при слабом освещении увеличивается чувствительность глаза. Для нейросуперпозиционных фасеточных глаз характерна суммация сигналов от зрительных клеток, находящихся в разных омматидиях, но получающих свет из одной и той же точки пространства. У некоторых насекомых (богомолы , поденки) одна часть глаза может быть построена по аппозиционному типу, а другая — по суперпозиционному.
В фасеточных глазах всех типов собственно светочувствительным элементом служат рабдомеры зрительных клеток, содержащие фотопигмент (обычно подобный родопсину). Поглощение фотопигментом квантов света — первое звено в цепи процессов, в результате которых зрительная клетка генерирует нервный сигнал.
Нервная проекция сетчатки на оптические ганглии мозга и, отчасти, особенности оптики фасеточных глаз таковы, что они обеспечивают анализ внешнего мира с точностью до растра омматидиев, а не отдельных зрительных клеток. Низкая угловая плотность омматидиев (их оптические оси расходятся под углами 1-6°) препятствует различению мелких деталей, однако малая инерционность в сочетании с высокой контрастной чувствительностью (1-5%) Ф. г. позволяет некоторым насекомым различать мелькания (мигания) света с частотой вплоть до 250-300 гц (для человека предельная частота около 50 гц). Фасеточные глаза обеспечивают многим беспозвоночным цветовое зрение с восприятием ультрафиолетовых лучей, а также анализ направления плоскости линейно-поляризованного света.
Почему членистоногие и особенно насекомые являются одной из наиболее процветающих групп живых существ?
Каковы преимущества и недостатки наружного скелета?
Какие функции выполняют многочисленные конечности у рака?
Почему насекомые больше полагаются на инстинктивное поведение, чем на приобретение навыков?
Тип Arthropoda (членистоногие) представлен большим количеством видов, живущих в настоящее время, чем все другие типы, вместе взятые. Известно более миллиона видов, составляющих такие процветающие классы, как ракообразные, паукообразные и самый крупный из всех — насекомые. На каждого человека приходится приблизительно 300 млн. насекомых. Палеонтологи говорят, что на Земле существовали эпохи пресмыкающихся и млекопитающих. Вероятно, без преувеличения можно сказать, что сейчас наступила эпоха насекомых.
Рассматривая тип в целом, можно утверждать, что на земном шаре нет места, которое не было бы занято одним из его представителей. Членистоногие обитают в океанских глубинах, в мелководных заливах и озерах, в сырых пещерах, в раскаленных пустынях, во влажных джунглях и в полярных зонах. Членистоногие могут плавать, летать, прыгать, ползать, извиваться или, подобно усоногим ракам, стоять на голове в известковом домике и забрасывать пищу в рот (табл. 5).
| Тип | Приблизительное число видов животных (Х1000) |
| Губки | 5 |
| Кишечнополостные | 5 |
| Плоские черви | 13 |
| Круглые черви | 10 |
| Кольчатые черви | 9 |
| Моллюски | 110 |
| — | Насекомые 675 |
| — | Ракообразные 25 |
| Членистоногие | 900 |
| — | Паукообразные 31 |
| — | Многоножки 2 |
| — | Двупарноногие 7 |
| Иглокожие | 6 |
| — | Рыбы 25 |
| — | Амфибии 3 |
| Хордовые | 45 |
| — | Рептилии 10 |
| — | Птицы 9 |
| — | Млекопитающие 4 |
| «Мелкие типы» | 20 |
В чем причина процветания типа членистоногих? Очевидно, в том, что их конечности имеют суставы с мышцами-антагонистами (противоположно действующими). Это свойство, которым, кроме них, обладают еще только позвоночные, обеспечивает членистоногих высокой подвижностью и делает их грозными охотниками. Кроме того, они имеют твердый наружный покров, или наружный скелет. Такой покров защищает их тело от высыхания и от нападения потенциальных врагов, однако несколько ограничивает свободу движений, и, наконец, насекомые были первой крупной группой животных, обитавших в лесах каменноугольного периода более 340 млн. лет назад. Почти полное отсутствие конкуренции привело к возникновению их большого разнообразия и позволило этим животным занимать почти любую экологическую нишу. Впоследствии (около 250 млн. лет назад) они приобрели и способность летать, что еще больше расширило занимаемую ими территорию. Различные виды членистоногих представлены на рисунке 15 1.
15.1. Некоторые примитивные членистоногие, такие, как трилобиты, вымерли, но дошедшие до нас ископаемые остатки свидетельствуют об их огромной численности
Наиболее примитивными из животных, которых можно отнести к членистоногим, были трилобиты (рис. 15-2). Они существовали 250-500 млн. лет назад и обитали в море. Эти животные имели тяжелые раковины, сегменты их тела и конечности были простыми и многократно повторяющимися. Эвриптериды, или морские скорпионы, достигавшие иногда в длину 2,5 м, жили, а затем исчезли примерно в то же время, что и трилобиты.

Одна древняя, относительно примитивная группа животных вымерла не полностью. Живущими представителями этого класса Xiphosura (мечехвостов) являются Limulus — подковообразные крабы (рис. 15-3). Эти необычные животные широко распространены вдоль Атлантического побережья США и в некоторых других районах земного шара. Зона их обитания — морское дно, где они охотятся за червями и другими организмами. Подковообразный краб, имеющий пять пар конечностей, очень похож на ископаемого подковообразного краба, вымершего 500 млн. лет назад.

При сравнении примитивных членистоногих с современными хорошо прослеживаются эволюционные изменения, произошедшие за это время. Конечностей стало меньше, многие из них либо трансформировались в клешни и части рта, либо стали трубками для переноса спермы; область головы стала зоной расположения органов чувств и нервной ткани; число сегментов уменьшилось, и они соединились, образовав голову, грудь и живот или комбинации из них.
15.2. Класс ракообразных — единственная многочисленная группа членистоногих, обитающих главным образом в воде
В этот крупный класс, состоящий из 28 000 видов, входит ряд хорошо известных животных: креветки, омары, крабы, раки и усоногие раки; менее известные организмы: равноногие раки, или мокрицы, и песчаные блошки; микроскопические веслоногие ракообразные и родственные им виды, которые составляют основную часть зоопланктона.
Веслоногие ракообразные — наиболее распространенные животные на Земле. Интересными представителями этих микроскопических или полумикроскопических ракообразных являются Daphnia и Leptodora (рис. 15-4). Leptodora обитает в чистых северных озерах. Почти полная прозрачность этого животного позволяет рассмотреть под микроскопом каждую его клетку. В головной части находится один сложный глаз, соединенный с мозгом длинным глазным нервом. Глаз производит сканирующие, или дрожательные, движения за счет быстрого сокращения мышц. Под микроскопом можно увидеть, как мышцы осуществляют движение туловищных отростков, сердцебиение и перистальтические сокращения кишечника. Leptodora имеет открытую кровеносную систему, в ее крови содержатся амебоподобные клетки. Питаться Leptodora может фитопланктоном или более мелкими животными, например дафниями.

15.3. Пресноводный рак — типичный представитель ракообразных
Рак Cambarus — широко распространенное ракообразное, обитающее в реках и озерах. Анатомия и физиология раков и омаров почти полностью идентична. Наиболее характерной особенностью обычного рака является большое количество туловищных отростков (рис. 15-5). Каждая пара отростков, хотя это и не видно с первого взгляда, отходит от определенного сегмента, или сомита, тела животного. Сегментация у раков выражена не так ярко, как, например, у кольчатых червей. Частично это связано с изменениями, которые выражаются в слиянии одних сегментов и перекомпоновке других.

У примитивных раков, вероятно, было много одинаковых парных отростков, которые в процессе эволюции развивались в разнообразные структуры. Cambarus имеет 18 пар отростков, расположенных на брюшной стороне тела, и одну слившуюся пару на хвосте. Все отростки снабжены суставами, а большинство из них состоит из наружной и внутренней ветвей. Оболочка, покрывающая отростки и тело рака, является твердым наружным скелетом или интегументом . Наружный скелет ракообразных более плотный и мощный, чем у насекомых и пауков, поскольку содержит соли кальция. Избыточная масса не имеет особого значения для животных, обитающих в воде, в отличие от наземных форм, масса которых ограничивает их размеры.
15.4. Интегумент заменяется по мере роста животного
Твердый наружный скелет, который выполняет защитную функцию, имеет два основных недостатка: он не может расширяться по мере роста животного и ограничивает сги бательные дви жения, которые важны для передвижения. Однако многочисленные суставы наружного скелета и сложная мускулатура частично компенсируют этот второй недостаток. Если же панцирь начинает сдерживать их рост, раки и другие членистоногие линяют, т. е. периодически сбрасывают наружный скелет.
Как только животное заполнит телом наружный скелет, под ним начинается образование нового, относительно мягкого панциря. В определенное время старый внешний скелет разрывается и рак выползает наружу, а новый интегумент начинает быстро твердеть. Рак, только что завершивший линьку, впитывает большое количество воды и набухает. После того как новый скелет затвердеет, рак теряет часть воды и уменьшается в размерах, что позволяет ему расти в пределах этого панциря. Процесс линьки повторяется обычно дважды в год в течение всей жизни животного (несколько десятилетий у определенных видов ракообразных).
В течение нескольких дней после линьки, до того как новый наружный скелет затвердеет, рак уязвим для хищников. Его мягкий интегумент легко мог^т разгрызть другие животные. В это времяу рак не способен даже обороняться, по-4 скольку его панцирь, ротовые части и конечности не укреплены защитной оболочкой. Кроме того, временно утрачивается часть пищеварительной системы, а также теряется способность поддержания равновесия в воде. Поэтому рак не может в этот период питаться и ориентироваться. Все, вместе взятое, делает процесс линьки опасным периодом для ракообразных. Между периодами линьки, когда рак имеет твердый панцирь, значительным преимуществом для него становится способность сбрасывать конечность или клешню, поврежденные хищником. Такой процесс называется аутотомией (самоампутацией). Отделение пораженного участка происходит у основания отростка, а потерю крови, которая может сопровождать этот процесс, предотвращают расположенные здесь диафрагма и клапан. Конечности у рака вновь образуются из близлежащих тканей. Во время следующей линьки новая конечность вырастает до размеров утраченной и способна выполнять присущую ей функцию. У омаров утраченная клешня восстанавливается сходным образом, но у крупных животных проходят годы, прежде чем она достигнет нужных размеров.
У рака хорошо развиты мышцы, которые, действуя независимо или совместно, осуществляют движение отдельных частей его тела. Большинство мышц прикреплено одним концом к неподвижному наружному скелету, а другим — к подвижной части тела, например к конечности. Брюшко рака является единственной гибкой частью тела, в которой мышцы располагаются по сегментам. У раков и омаров эта часть тела наиболее мясистая.
Все мышцы группируются в антагонистические пары и обычно обозначаются как экстензоры (разгибатели) и флексоры (сгибатели). Далее механизм аутотомии снабжен собственным набором мышц, которые при определенных условиях обеспечивают сбрасывание конечности.
15.5. Ракообразные и насекомые имеют сложные фасеточные глаза, которые позволяют им хорошо ориентироваться и обнаруживать движущиеся объекты
Сложные, фасеточные глаза значительно отличаются от обычных глаз, разделенных на камеры, например у млекопитающих (рис. 15-6). Каждый фасеточный глаз состоит из множества мелких воспринимающих свет фасеток, называемых омматидиями. Нa внешнем крае этих вытянутых структур расположены тончайшие линзы. Каждая фасетка имеет собственную иннервацию, а в одном фасеточном глазе содержится около 10 000 омматидиев.

Оси каждой из фоторецепторных единиц расположены веерообразно, под небольшим углом друг к другу и воспринимают источник света, чаще солнце, каждый в своей немного отличающейся плоскости. Когда объект находится между источником света и глазом, вся информация от отдельных единиц объединяется и создается определенная картина объекта. Такой глаз может обнаруживать двигающийся в любом направлении объект поблизости от животного. Поскольку членистоногие имеют два глаза, они могут оценивать и расстояние до объекта.
15.6. Омары, морские «родственники» речных раков, образуют химическое вещество, которое определяет их брачное поведение
Самки омаров обычно вступают в период размножения сразу после линьки. Недавно было установлено, что перед спариванием они выделяют феромон, крустэкдизон , который изменяет поведение самца омара. Известно много таких веществ, действующих сходным образом у насекомых и млекопитающих. Самец омаров очень агрессивен и в обычных условиях сразу атакует особь любого пола, которая помещена с ним в один аквариум. Однако феромон половозрелой самки уменьшает агрессивность самца, поэтому он может спариваться с прошедшей линьку самкой, не причиняя ей повреждений. Самец приближается к самке, переворачивает ее на спину и откладывает сперму около полового отверстия. После этого поведенческие реакции, связанные с размножением, полностью прекращаются.
15.7. Ни одна группа организмов не может сравниться по разнообразию с классом насекомых
Насекомые представляют собой чрезвычайно разнообразную и распространенную группу животных, адаптированных к почти любой экологической нише земного шара, за исключением океанов.
Несмотря на разнообразие насекомых, они имеют ряд общих свойств. Все они имеют несколько видов твердого, водонепроницаемого наружного скелета и систему трахеального дыхания, в которой используются мелкие трубочки, несущие воздух непосредственно к клеткам. Тело насекомых состоит из трех основных частей — головы, грудных сегментов и брюшка; насекомые также имеют одну пару антенн (у ракообразных — две пары), пару фасеточных глаз, три ротовые части, которые представляют собой модифицированные конечности, и три пары ног, прикрепленных к грудным сегментам.
Энтомологи различают около миллиона современных насекомых, входящих в 26 отрядов, но их можно более просто разделить соответственно уровням, которые они занимают на эволюционной лестнице (рис. 15-7).

Большинство примитивных насекомых — бескрылые формы, и их называют аптериготами , например шетинкохвостки. Все остальные относятся к птериготам — крылатым насекомым, которые, в свою очередь, можно подразделить на насекомых, неспособных складывать крылья за спиной, например стрекозы, и насекомых, у которых крылья компактно складываются, например у жуков, мотыльков, кузнечиков, мух, пчел, муравьев и т. д. Насекомых, которые могут складывать крылья, иногда классифицируют по типу их развития.
Некоторые насекомые появляются из яйца в виде форм, сходных со взрослыми; другие развиваются из яйца в личиночную форму, непохожую на взрослый организм, например, гусеницы. Эти различия часто означают, что личинки специализированы к усиленному питанию, взрослые же формы специализированы к размножению.
Даже такое краткое описание разнообразия насекомых позволит читателю оценить, насколько сложен выбор одного типичного представителя из огромного числа видов насекомых. Это сделать невозможно.
Поэтому было выбрано наиболее знакомое нам и доступное насекомое — комнатная муха.
15.8. На распространение мух повлияло увеличение численности человечества в современном мире
С ростом численности населения на земном шаре увеличивается количество отходов, которые служат источником пищи для мух и их личинок. Именно этим объясняется такое огромное распространение мух на Земле.
Подобно другим насекомым, комнатная муха имеет плотную хитиновую оболочку, которая защищает ее внутренние органы и ткани от высыхания. Конечности у Musca очень отличаются от конечностей рака, хотя у тех и других они состоят из члеников. Ротовые выросты у мухи приспособлены для всасывания пищи, конечности используются для передвижения. Пара крупных летательных крыльев расположена на грудной части тела, вторая пара сильно редуцированных крыльев действует как органы равновесия. Большинство же других насекомых имеют две пары летательных крыльев.
Внутреннее строение насекомых на первый взгляд просто. Имеется пищеварительная трубка со специализированными участками пищеварения и всасывания, мужские или женские гонады, выделительные органы, которые опорожняются в кишку, открытая сосудистая система, удвоенный брюшной нервный ствол, различные железы, специальные запасающие ткани, и сильные мышцы, особенно в области крыльев. Кроме того, имеется трахейная дыхательная система, которая состоит из заполненных воздухом трубок, проникающих во все ткани тела насекомого.
Насекомые являются единственной группой летающих беспозвоночных, что частично обусловливается их своеобразной дыхательной системой. На внешней оболочке насекомых расположены наружные отверстия выстланных хитином трубочек, которые, простираясь внутрь, разветвляются на все более мелкие трубки и доставляют кислород к отдельным клеткам или небольшим группам клеток. Углекислый газ, образующийся в тканях, выводится через те же дыхательные трубки. Таким образом, насекомые имеют наиболее эффективную дыхательную систему, которая способна быстро обеспечивать кислородом такие метаболически активные ткани, как летательные мышцы.
Маленькие клапаны, или дыхальца, расположенные у наружных отверстий дыхательных трубок, регулируют подачу и удаление воздуха. Поскольку трубки заполнены водяными парами, дыхальца также предотвращают потерю воды. Хитин в дыхательной трубке расположен спирально, что позволяет трубкам растягиваться линейно при движении насекомого и в то же время предотвращает перегибание и сильное сжатие внутреннего просвета этих тонкостенных трубок. Кроме эффективного переноса газов, этот тип дыхательной системы имеет и другие преимущества. Например, такая малая часть системы, как дыхальца, противостоит обезвоживающему действию окружающей среды и обеспечивает сохранение воды в теле насекомого.
15.9. С помощью конечностей муха способна ощущать вкус пищи и удерживать тело на вертикальной поверхности
Муха засасывает пищу, выдвигая длинный трубчатый хоботок, который связан со ртом, однако этот орган не позволяет ей ощущать вкус пищи. Только недавно было установлено, что эту функцию выполняют конечности животного. На передних ножках мухи расположены маленькие волоски, которые связаны нервами с другими частями тела, а главным образом с хоботком. Если муха натолкнется на субстрат, который ощущают ее конечности, а нервная система устанавливает, что он съедобен, животное выдвигает хоботок и начинает питаться.
Кроме этих чувствительных волосков, на концах конечностей мухи расположены маленькие подушечки, которые содержат железы, образующие небольшое количество липкого вещества. С помощью этого вещества легкое насекомое прикрепляется к стене, потолку или скользкой поверхности ведра для отбросов. Конечности некоторых насекомых снабжены различными шипами, клешнями и выступами, которые позволяют им прикрепляться к растениям и животным.
15.10. Небольшие размеры насекомых и устоявшиеся поведенческие акты способствуют их успеху в эволюции
Во вступлении к этой главе был приведен ряд соображений, объясняющих процветание членистоногих, в частности членистый наружный скелет, а по отношению к насекомым — способность летать. Кроме того, малые размеры насекомых и особый тип нервной системы, очевидно, обеспечивают преимущество над конкурирующими с ними животными. Мелкие животные могут занимать такие места обитания, которые недоступны для остальных животных. Другие наземные формы, подавляющее большинство которых относится к позвоночным, обычно более крупные и не могут жить в узких расщелинах, под скалами или в трещинах на стволах деревьев. Поэтому конкуренция за территорию у мелких наземных животных выражена значительно слабее, хотя некоторые виды насекомых вступают в конкурентные отношения с другими насекомыми за одну территорию.
Еще одно очевидное эволюционное преимущество, связанное с уменьшением размеров организма, включает изменения величины и силы мышц. Муравьи, осы и другие насекомые способны поднимать намного большую массу по отношению к их размерам, чем, например, крыса, слон или человек. Первоклассные штангисты могут поднять штангу, равную их собственной массе, не более 2-3 раз, в то время как обычный муравей способен поднять 10-15 раз груз, равный его массе, и перенести его на значительное расстояние. Это происходит не за счет особой силы мышц муравья, а является результатом специфического соотношения между размером и силой мышц. И наконец, еще одним свойством насекомых, которое дает им значительное эволюционное преимущество, является характер их поведения. Поведение в этом случае определяется как результат функции нервной системы, который обычно реализуется в двигательной активности насекомых. Из-за отсутствия лучшего термина поведение насекомого обозначается как инстинктивное. Следует со всей отчетливостью констатировать, что в настоящее время не существует ни одного точного объяснения первых процессов, лежащих в основе данного типа поведения, но мы можем по крайней мере охарактеризовать эти поведенческие реакции.
Инстинктивное , или врожденное поведение базируется на комплексе наследственных или устоявшихся особенностей реакций, которые приводятся в действие определенными стимулами. Многократно утверждалось, что обучение и полученный прежде опыт не требуются для данного типа поведения, однако и то и другое все-таки играет определенную роль в ряде инстинктивных поведенческих актов. Примером инстинктивного поведения является описанное ранее поведение самки кальмара, которая в процессе откладывания яйцеклеток прикрепляет их к скалам. Известно много примеров подобного поведения также и среди членистоногих. Размер и форма гнезд, которые строят насекомые, определяются ими инстинктивно. Насекомые могут строить одно гнездо, не ориентируясь на построенное ранее. Самцы некоторых видов москитов летят на звучащий камертон, настроенный в тон со звуком, издаваемым самкой, даже если они никогда раньше не слышали подобного звука. Но самцы не отвечают на звучание камертона, если его тон даже незначительно отличается от звука самки.
Впечатляющий пример инстинктивного поведения касается размножения богомола. Самец богомола, находящийся рядом с самкой, не вступает в копуляцию до тех пор, пока самка в начальной фазе размножения не откусит ему голову. Очевидно в мозге самца имеются тормозящие центры, которые не позволяют копуляторным нервам активизировать другие части тела к размножению. И действительно, неполовозрелые самцы после декапитации пытаются копулировать с самкой. Тот факт, что самец богомола может спариваться с самкой, даже когда у него удалена голова и мозг и эта его активность не связана с обучением, позволяет предположить сложные механизмы, заложенные в таких инстинктивных действиях.
Нервная система, обусловливающая инстинктивное поведение, несомненно, оберегает животное от возможных ошибок и затрат времени и энергии на усвоение опыта. Значительно проще следовать установленным правилам или инструкциям в ответ на точные стимулы, указывающие последовательность действий. У насекомых, имеющих короткий период жизни, нет времени на обучение. В течение их эволюционной истории способность корректировать или менять поведенческие реакции при возникновении неожиданных опасностей сопровождалась крупными потерями. Однако их процветание, связанное с инстинктивным поведением, говорит в пользу большей его важности по сравнению с поведением, основанным на обучении у животных сходного уровня сложности.
В процессе эволюции зрения у некоторых животных возникают довольно сложные оптические приборы. К таким, безусловно, можно отнести глаза фасеточные. Они сформировались у насекомых и ракообразных, некоторых членистоногих и беспозвоночных. Чем отличается фасеточный глаз от простого, каковы его основные функции? Об этом поговорим в нашем сегодняшнем материале.
Глаза фасеточные
Это оптическая система, растровая, где отсутствует единая сетчатка. А все рецепторы объединены в небольшие ретинулы (группы), образуя выпуклый слой, не содержащий более никаких нервных окончаний. Таким образом, глаз состоит из множества отдельных единиц — омматидий, объединяющихся в общую систему зрения.
Глаза фасеточные, присущие, к отличаются от бинокулярных (присущих в том числе и человеку) плохим определением мелких деталей. Зато они способны различать колебания света (до 300 Гц), тогда как для человека предельные возможности — 50 Гц. А еще мембрана такого типа глаз имеет трубчатую структуру. Ввиду этого глаза фасеточные не имеют таких особенностей рефракции, как дальнозоркость или близорукость, для них неприменимо понятие аккомодации.

Некоторые особенности строения и зрения
У многих насекомых занимают большую часть головы и фактически неподвижны. К примеру, глаза фасеточные у стрекозы состоят из 30 000 частиц, образуя сложную структуру. У бабочек — 17 000 омматидиев, у мухи — 4 тысячи, у пчелы — 5. Наименьшее количество частичек у муравья рабочего — 100 штук.
Бинокулярное или фасеточное?
Первый тип зрения позволяет воспринимать объем предметов, их мелкие детали, оценивать расстояния до объектов и их расположение по отношению друг к другу. Однако человека ограничивается углом в 45 градусов. Если обзор необходим более полный, глазное яблоко осуществляет движение на рефлекторном уровне (либо мы повернем голову вокруг оси). Фасеточные глаза в виде полусфер с омматидиями позволяют видеть окружающую действительность со всех сторон, не поворачивая органов зрения или головы. Причем изображение, которое передает при этом глаз, очень похоже на мозаику: одной структурной единицей глаза воспринимается отдельный элемент, а вместе они отвечают за воссоздание полной картины.

Разновидности
Омматидии имеют анатомические особенности, в результате чего и различаются их оптические свойства (к примеру, у разных насекомых). Ученые определяют три вида фасета:

Кстати, некоторые виды насекомых имеют смешанный тип фасеточных органов зрения, а у многих, кроме рассматриваемых нами, имеются еще и простые глаза. Так, у мухи, к примеру, по бокам головы расположены парные фасеточные органы довольно больших размеров. А на темени есть три простых глаза, выполняющих вспомогательные функции. Такая же организация органов зрения и у пчелы — то есть всего пять глаз!
У некоторых ракообразных фасеточные глаза как бы сидят на подвижных выростах-стебельках.
А у некоторых амфибий и рыб имеется еще и дополнительный (теменной) глаз, который различает свет, но обладает предметным зрением. Сетчатка его состоит только из клеток и рецепторов.
Современные научные разработки
В последнее время глаза фасеточные — предмет изучения и восторга ученых. Ведь такие органы зрения, ввиду своего оригинального строения, дают почву для научных изобретений и изысканий в мире современной оптики. Основные преимущества — широкий обзор пространства, разработка искусственных фасеток, используемых преимущественно в миниатюрных, компактных, секретных системах наблюдения.
Основные признаки членистоногих:
Гетерономная сегментация тела, т.е. разные участки тела обладают сегментами разного строения, причем многие сегменты сливаются друг с другом. Тело разделено на голову, грудь и брюшко (у скорпионов — два брюшка). У паукообразных и некоторых ракообразных голова и грудь объединяются в головогрудь;
Членистые конечности разделены на отделы, которые соединяются друг с другом и с телом суставами. Конечности, расположенные на разных отделах тела, дифференцированы по функциям — захват пищи, передвижение, дыхание и др.;
Тело покрыто хитиновой кутикулой, состоящей из липидов, протеинов, хитина, углекислой извести. Растут членистоногие, только сбросив хитиновый покров во время линьки;
Мускулатура представлена отдельными мышечными пучками; мышечные волокна имеют поперечно-полосатую структуру;
Полость тела (миксоцель) смешанная, образованная из первичной полости и слившихся целомических мешков. В ней располагаются внутренние органы;
Пищеварительная система состоит из передней, средней и задней кишок; пища обрабатывается секретами пищеварительных желез;
Кровеносная система незамкнутая, имеется сердце. В кровеносной системе циркулирует гемолимфа, состав которой частично соответствует составу крови, а частично — составу целомической жидкости;
Органы дыхания представлены жабрами, легкими и трахеями;
Нервная система по строению сходна с нервной системой кольчатых червей. Происходит еще большее слияние ганглиев брюшной нервной цепочки;
Выделительная система представлена мальпигиевыми сосудами;
Размножаются только половым путем, обычно раздельнополы, часто выражен половой диморфизм. Развитие как прямое, так и непрямое.
Наиболее важны классы: ракообразные, паукообразные и насекомые.
Ракообразные представлены в основном водными животными. Некоторые, например крабы имокрицы, могут жить на суше. Низшие ракообразные: дафнии, циклопы. Высшие ракообразные:раки, омары, креветки, крабы и др.
Покровы животных образованы хитиновой кутикулой, образующей наружный скелет.Мышцы образуют пучки, соединяющие соседние сегменты или членики конечности.
На голове речного рака находятся пять пар придатков. Первая пара — антеннулы, выполняющие функции органов осязания и обоняния. Вторая пара — антенны, имеющие вид длинных усиков и выполняющие функции органов чувств, а иногда и передвижения. Затем следуют верхние челюсти — жвалы (мандибулы), размельчающие пищу, и первые и вторые нижние челюсти — максиллы. Первые три пары грудных конечностей (ногочелюсти) речного рака выполняют функции дыхания (жабры), захвата, удержания, измельчения и проталкивания пищи. Следующая пара грудных конечностей образована мощными клешнями. Ходильные ноги имеют одноветвистое строение. В их основании расположены жабры. Брюшные ноги развиты слабо и выполняют дыхательную функцию, а у самцов первые две пары брюшных ножек видоизменены в совокупительный аппарат.
Пищеварительная система состоит из трех отделов. Кишка хитинизирована, что важно для перетирания пищи. Желудок разделен на жевательный и цедильный отделы. В жевательном отделе кутикула толще и имеет зазубрины. В цедильной части пища фильтруется. Есть пищеварительная железа, проток которой открывается в короткую среднюю кишку. Она совмещает функции печени и поджелудочной железы. Задняя кишка заканчивается анальным отверстием.
Дыхание. Раки дышат жабрами, расположенными на конечностях. Органы дыхания у водных животных — жабры, у наземных — легкие или трахеи. Жабры и легкие — видоизмененные конечности, трахеи (ветвящиеся трубочки) образуются в результате впячивания покровов.
Кровеносная система животных не замкнута. Она состоит из сердца, расположенного на спинной стороне тела, и крупных сосудов. Гемолимфа из сердца поступает в сосуды, а из сосудов в полость тела, затем она направляется к жабрам, где происходит насыщение гемолимфы кислородом, после чего она снова поступает в сердце.
Нервная система раков близка по строению к нервной системе кольчатых червей. Развитие идет в направлении сближения нервных узлов. Достаточно сложно устроен их головной мозг. Нервная система состоит из головного нервного узла («головной мозг»), окологлоточного кольца и брюшной нервной цепочки. Часто наблюдается слияние ганглиев брюшной нервной цепочки с образованием крупных нервных узлов. Высокого развития достигают все органы чувств: осязания, обоняния, вкуса, зрения, слуха и равновесия
Выделительная система представлена двумя парами выделительных (зеленых) желез. Их протоки открываются у основания антенн (первая пара) и у основания второй пары нижних челюстей (вторая пара). Извитые каналы желез впадают в мочевой пузырь.
Размножение. Раки раздельнополы. Развитие речного рака происходит без метаморфоза. Его первая и вторая пара брюшных ножек превратились в копулятивные органы. Сперматозоиды проникают в семяприемники и там хранятся до момента выхода яиц из женских половых отверстий. Тогда и наступает оплодотворение. Из яйца выходит маленький рачок.
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODА)
Среди всех групп беспозвоночных животных тип членистоногих выделяется наибольшим разнообразием приспособлений к самым различным условиям существования, изумительным богатством форм и огромным числом видов. Число видов членистоногих приближается к 3 миллионам и намного превышает число видов всех остальных типов животных и растений, вместе взятых, включая микроорганизмы. Особенно обильны видами насекомые, на долю которых падает более 90% известных видов членистоногих.
Именно тонкими приспособлениями к использованию всевозможных местообитаний, всяких источников ппщи, часто недоступных для других животных, объясняется многообразие и обилие видов членистоногих.
Трудно найти такие места, где не было бы членистоногих. Они живут во всех морях и океанах — и в толще воды, и на дне, и в грунте на разных глубинах; они обитают и во всевозможных пресных водоемах — не только в реках и озерах, но и в мелких пересыхающих лужах, в подземных водах и пещерах, в скоплениях воды в дуплах деревьев. Некоторые виды приспособились к жизни в очень соленых водоемах, другие — в горячих источниках. Кажется, все разнообразие водной среды на поверхности нашей планеты освоено членистоногими.
Большая армия представителей этого типа приспособилась и к жизни вне воды — на суше, где членистоногие особенно многообразны. Весь слой почвы, пронизанный корнями растений, обильно заселен и членистоногими — во многих лесных и луговых почвах миллион особей клещей и ногохвосток на 1 м2 не редкость. Огромное количество членистоногих обитает на поверхности различных частей растений и внутри растительных тканей.
Во всех климатических зонах можно встретить членистоногих. Огромно их количество и велико многообразие в тропиках, но есть они и в высоких широтах Арктики и на побережье Антарктиды, где недавно были обнаружены новые виды клещей и ногохвосток. Членистоногих можно встретить и в самых сухих пустынях. Образно можно сказать, что членистоногие — это тип животных, который по полноте овладения поверхностью земного шара не имеет равных.
По многим признакам строения членистоногие близки к кольчатым червям, особенно к полихетам. Замечательный зоолог начала прошлого века Жорж Кювье, предложивший деление животных на типы, объединил кольчатых червей и членистоногих в единый тип членистых животных (Articulata). Знаменательно, что и в наши дни некоторые авторитетные зоологи, например В. Н. Беклемишев, ратуют за восстановление типа членистых. Это показывает, что членистоногие во всяком случае очень близки к многощетинковым кольчатым червям. От близких к ним форм когда-то давно, еще в те геологические эпохи, в отложениях которых не сохранилось ископаемых остатков, отделились предки современных членистоногих.
Членистоногие, как и кольчатые черви, — это двустороннесимметричные сегментированные животные. Сходство членистоногих с кольчецами проявляется не только в наружной сегментации. Центральная нервная система у более примитивных членистоногих очень сходна с лестничной нервной системой полихет.
Специфические признаки членистоногих связаны с тем, что у них тело покрыто очень плотной и прочной кутикулой, которая нередко образует очень твердый панцирь. Кутикула у членистоногих вы деляется наружным слоем клеток, так называемой гиподермой, имеет очень сложное строение и сложный химический состав. Раньше считали, что у членистоногих кутикула состоит только из хитина. Теперь выяснилось, что, кроме хитина, в состав ее входят многие другие соединения, в основном задубленные белки, и что у большинства членистоногих на долю хитина падает не больше трети веществ, из которых состоит кутикула. Хитин — это сложный полисахарид, в состав молекулы которого входят атомы азота. Вероятно, это не определенное соединение, а группа близких соединений, формулы которых точно не установлены. Хитин эластичен, прочен и устойчив к химическим воздействиям. Нередко в кутикуле содержится много углекислого кальция.
Прочность и твердость кутикулы членистоногих настолько велики, что в тех местах, где она достигает большой толщины, покровы становятся негибкими. Подвижность сегментов тела друг относительно друга, обеспечивающая движение, у членистоногих достигается тем, что между твердыми участками (склеритами) сохраняются перепончатые участки с очень тонкой кутикулой; такие перепонки на сгибах образуют складки, расправляющиеся при растягивании тела. Далеко не все сегменты у членистоногих подвижны друг относительно друга, очень часто наблюдается полное слияние соседних сегментов, особенно в передних отделах тела.
В теле членистоногих обычно можно различить 3 отдела — голову, на которой находятся основные органы чувств, служащие для ориентировки в пространстве (глаза, усики и др.)» и органы захватывания пищи. Сегменты головы у членистоногих слитные. Далее следует грудь — тот отдел тела, на котором располагаются основные двигательные придатки — ноги, а у насекомых и крылья. Задний отдел тела называется брюшком. У некоторых членистоногих (например, у многоножек, а из ископаемых — у трилобитов) все туловищные сегменты более или менее однородны, и тогда в теле животного различают только 2 отдела — голову и туловище. У многих членистоногих (у паукообразных, десятиногих раков) сливаются головной и грудной отделы, образуя головогрудь. А у клещей вообще утрачиваются границы между отделами тела, тело слитное.
Передвигаются членистоногие с помощью конечностей.
Поскольку конечности у них, как и все тело, покрыты кутикулой, их движение возможно тоже только при разделении ног на членики. По строению конечностей, на которые опираются членистоногие при движении по твердой поверхности или с помощью которых они плавают, весь тип и получил свое название.
Некоторое количество пар конечностей на голове у всех членистоногих превращается в ротовые органы — челюсти, использующиеся хотя бы частично для захватывания или размельчения пищи. Это тоже признак, характерный для типа членистоногих. Ротовые части бывают настолько видоизменены, что их трудно даже признать конечностями, например, хоботок комара или клопа. Число пар ног, которые служат для передвижения, неодинаково у разных членистоногих. Для насекомых характерно развитие 3 пар двигательных конечностей, для паукообразных — 4 пар, а у некоторых многоножек число ног превышает сотню.
Нередко туловищные конечности служат не только как ноги, но и как жабры, присоски и другие органы.
У членистоногих двигательная мускулатура состоит из отдельных мышц, концы которых присоединяются к разным утолщенным участкам покровов, отделенным друг от друга мягкой перепонкой. Движение членистоногих обеспечивается не изгибами тела при сокращении всей мускулатуры туловища, как у червей, а сокращениями обычно немногих мышц, связанных с конечностями или другими двигательными придатками (крылья насекомых, вилочка ногохвосток и др.)- Мышцы членистоногих обладают исключительно высокой способностью к сокращению. Все они поперечнополосатые, причем обнаруживаемая под микроскопом поперечная исчерченность мышечных волоконец у членистоногих выражена больше, чем у позвоночных, — мышцы членистоногих сильнее наших, если, конеч но, учитывать размеры. Так как панцирь членистоногих служит и местом прикрепления двигательных мышц, он является и скелетом, называемым «наружным» в отличие от привычного нам внутреннего скелета позвоночных.
Панцирь членистоногих лишь в результате долгой эволюции стал скелетом. А сначала утолщение и упрочнение покровов у обитавших в морях далеких червеобразных предков членистоногих служило, надо полагать, приспособлением к защите внутренних органов от повреждений. Утолщение и уплотнение кутикулы делает ее менее проницаемой. Поэтому понятно, что только очень мелкие членистоногие, имеющие тонкие покровы, живущие в воде или в очень влажных местах, могут дышатьвсейповерхностыо тела.
У всех более крупных членистоногих, имеющих и более плотные покровы, обязательно развиваются специальные дыхательные органы, поверхность которых покрыта такой тонкой кутикулой, что через нее легко может поступать кислород.
У живущих в воде членистоногих это жабры. Для дыхания в воде важно, чтобы поверхность жабр была возможно большей. Поэтому жабры всегда представляют собой тонкие листовидные выросты или пучки пальцевидных отростков. Обычно жабрами у членистоногих становятся конечности или части конечностей. В воде благодаря большой ее плотности даже тоненькие жаберные листочки не слипаются друг с другом. Поэтому на небольшом участке тела (в остальных местах защищенного плотными покровами) может разместиться много листовидных жабр.
Для доставки кислорода от жабр ко всем удаленным от них органам служит кровеносная система и полостная жидкость. У членистоногих кровеносная система представлена расположенным в спинной части тела пульсирующим, обычно продолговатым сосудом — сердцем. Кровь поступает в сердце через закрывающиеся клапанами боковые отверстия, а выталкивается из него в полость тела или непосредственно, или через немногие крупные сосуды — система кровообращения незамкнутая. У крупных морских членистоногцх, имеющих толстый панцирь, — у омаров, у мечехвостов — в крови есть дыхательный пигмент гемоцианин, сходный по химическому составу с нашим гемоглобином, но придающий крови не красный, а синеватый цвет: в состав гемоцианина входит не железо, как в гемоглобине, а медь. Гемоцианин легко присоединяет кислород при прохождении крови через жабры и отдает его внутренним органам. Так как у членистоногих кровеносная система незамкнутая, правильнее называть их «кровь» «гемолимфой», но и термином «кровь» зоологи широко пользуются.
На открытой поверхности суши ни кожное дыхание, ни дыхание с помощью жабр невозможно легкопроницаемые участки покровов пропускают воду и быстро подсыхают, теряя проницаемость. Поэтому водные организмы, извлеченные на сушу, погибают от потери воды или от недостатка поступающего кислорода. На суше только скрытоживущие мелкие членистоногие, обитающие в таких местах, где воздух всегда насыщен водяным паром, например в почве, могут дышать всей поверхностью тела (мелкие клещи, ногохвостки, симфилы). Жизнь на суше вне влажных укрытий требует развития непроницаемых для испарения покровов, а такие покровы не пропускают и кислород.
У наземных членистоногих развиваются особые дыхательные органы — либо «легкие», представляющие глубокие мешковидные впячивания со складчатыми стенками, открывающиеся наружу лишь небольшими отверстиями (у пауков, у скорпионов), либо (у большинства наземных членистоногих) трахеи. Трахеи представляют собой тонкие, обычно обильно ветвящиеся трубочки, открывающиеся наружу маленьким входным отверстием («дыхальцем») на поверхности тела. Ветви этих трубочек омываются полостной жидкостью, а самые мелкие ответвления доходят до отдельных клеток. Из тонких трубочек трахей вода через дыхальца испаряется очень медленно, и потери ее незначительны, а кислород по ним проникает быстро и легко.
Уже далекие предки разных групп членистоногих приспособились к жизни в разных средах. Можно выделить группу водных членистоногих, к которым относятся ракообразные, выделяемую в подтип жабродышащих . Правда, среди современных ракообразных есть и такие, которые живут на суше (мокрпцы), но по всем признакам они очень близки к своим водным родичам. Другой подтип — трахейнодышащие — представлен типично наземными членистоногими — различными группами многоножек и насекомых. Они прекрасно приспособлены к жизни на суше. В их покровах содержатся воскои жироподобные вещества, защищающие организм от высыхания, а все системы органов, связанных с расходованием воды, — выделительная и дыхательная — действуют так, что влага расходуется минимально. Среди насекомых есть отдельные виды, проводящие всю жизнь или часть ее в воде, но строение таких водных насекомых явно показывает их происхождение от обитавших на суше предков.
За длившуюся сотни миллионов лет историю эти группы обособились в связи с приспособлениями к жизни в двух разных средах — в воде и на суше.
Третий подтип членистоногих — хелицеровые — очень существенно отличается и от ракообразных, и от трахейнодышащих. Среди хелицеровых есть и водные представители — мечехвосты, обитающие в морях, и наземные — паукообразные.
В ископаемом состоянии сохранилось много остатков хелицеровых. Их изучение, дополняющее изучение современных представителей, показывает, как шло разделение классов в результате приспособления к жизни в разных средах-в воде и на суше. Изучение всех групп членистоногих убеждает в том, что переход их к жизни на суше был связан со способностью к выработке предохраняющих от испарения влаги покровов, к сохранению в теле воды, без которой невозможна жизнь.
У членистоногих, хорошо приспособившихся к жизни на суше, панцирь бывает нередко тоньше, чем у обитающих в воде, но зато на его поверхности отлагается очень тонкий слой воскоподобных и жироподобных веществ, не пропускающих пары воды. Этот слой защищает организм от высыхания и препятствует испарению. Поэтому у всех членистоногих, живущих открыто на поверхности суши, есть специальные органы дыхания. Те членистоногие, у которых специальных органов дыхания нет, которые дышат всей поверхностью кожи, вынуждены обитать во влажных укрытиях — в почве, в лесной подстилке ит. д., только иногда по ночам и после сильных дождей показываясь на поверхности.
У наилучшим образом приспособившихся к жизни на суше членистоногих чаще всего развивается трахейная система. Не только дыхательная, но и другие системы органов меняются у членистоногих с переходом к жизни на суше.
Выведение растворенных вредных продуктов разрушения белков у водных членистоногих происходит непосредственно во внешнюю среду через отверстия выделительных органов, открывающиеся у основания конечностей головного или грудного отдела. А у всех членистоногих, наилучшим образом приспособившихся к жизни на суше, — у пауков, многоножек, насекомых — выделительные трубочки («мальпигиевы сосуды») открываются в начале задней кишки. По мере прохождения по ней непереваренных остатков пищи и продуктов выделения и из тех и из других стенки задней кишки всасывают воду, столь труднодоступную для жителей суши, а экскременты и продукты обмена выводятся из кишечника в обезвоженном состоянии.
Приспособившиеся к жизни на суше членистоногие выработали способность запасать много жира в особых образованиях- в жировом теле. Главная роль жирового тела состоит в том, чтобы в критические периоды, когда организм не может получить воду извне, снабжать его. водой!
При окислении жиров образуется много воды (так называемая «метаболическая вода»), которая и идет на покрытие расхода влаги животным. В этом отношении насекомые сходны с верблюдами, у которых при длительных переходах через пустыню используется вода, образующаяся при окислении запасов жира в горбу. У водных групп членистоногих (ракообразных, мечехвостов) жирового тела не бывает.
Наружный панцирь членистоногих настолько плотен, что в затвердевшем состоянии не растягивается. Поэтому рост членистоногих сопровождается линьками. У растущего животного старый панцирь становится тесным, лопается, обычно сперва на спине, и сбрасывается. И только в короткий период, пока новый панцирь, образовавшийся под ним, не затвердеет, животное может увеличиваться в размерах. Во время сбрасывания старых покровов, которые сходят не только со всей наружной поверхности тела, но и со всех конечностей, в том числе и с ротовых органов, а также с передней и задней кишки и со всех дыхательных органов, членистоногие бывают вялыми, малоподвижными, не питаются и стараются спрятаться в каких-либо укрытиях.
Рост у членистоногих — не непрерывный процесс, а ступенчатый, размеры тела увеличиваются скачкообразно — сразу после линьки. У многих членистоногих во время линьки происходит и большее или меньшее изменение строения тела. Часто взрослое животное приспособлено к совсем иному образу жизни, чем молодое, и потому особи разных возрастов (возрастом у членистоногих называется период между линьками) бывают даже совсем непохожи друг на друга. В таком случае развитие сопровождается метаморфозом. Превращение гусеницы в куколку, а куколки в бабочку может служить общеизвестным примером.
Образ жизни разных членистоногих неодинаков. Многие из них отличаются очень сложным поведением, что всегда связано с высоким развитием органов чувств и центральной нервной системы.

,


У большинства членистоногих хорошо развиты глаза. Глаза у членистоногих бывают простые, имеющие одну линзу, и сложные, или фасеточные. У высших представителей всех подтипов глаза сложные, фасеточные, состоящие из множества расположенных плотно рядом друг с друном маленьких глазков. Иногда число глазков (так называемых омматидиев) в фасеточном глазу измеряется несколькими тысячами (рис. 185). Фасеточные глаза характерны для высших ракообразных, мечехвостов и насекомых. Такие глаза были и у живших в палеозойскую эру трилобитов — членистоногих, полностью вымерших в конце этой эры. Каждый омматидий воспринимает только одну точку находящегося перед ним предмета (рис. 186). В результате сложный глаз дает изображение, состоящее из большого числа отдельных точек, т. е. мозаичное (как тоновое клише в книге) и прямое (рис. 187, см. т. 3, табл. 18). Вспомним, что изображение, получаемое в нашем глазу, перевернутое и цельное!

Глаза членистоногих, оказывается, могут воспринимать не только те лучи, которые видимы для нашего глаза, но и ультрафиолетовую часть спектра. Например, многие цветы, которые нам кажутся равномерно белыми, для насекомых оказываются пестрыми; у таких цветов есть не видимый нами ультрафиолетовый рисунок (см. т. 3, табл. 19).
И наземные и водные членистоногие способны различать обычный свет и так называемый поляризованный (те световые лучи, электромагнитные колебания которых совершаются в одной плоскости). Поскольку для рассеянного света неба характерна определенная степень поляризации лучей в меридиональной плоскости, членистоногие по небу, как по компасу, могут ориентироваться и в ясные и в пасмурные дни. Таким образом, членистоногие различают не только частоту, но и плоскость колебаний световых лучей.
Мы не ощущаем магнитные волны. А например, самки термитов в гнезде лежат так, что ось их тела направлена по магнитному меридиану, а если поместить рядом сильный магнит, меняют свое положение. Как воспринимают членистоногие магнитные волны, пока не выяснено. Отмечено, что членистоногие могут реагировать и на короткие радиоволны.
- http://fb.ru/article/250729/glaza-fasetochnyie-chem-otlichayutsya-ot-prostyih
- http://isee.by/sections/other/facet_vision.html
- http://gufo.me/dict/biology/%D1%84%D0%B0%D1%81%D0%B5%D1%82%D0%BE%D1%87%D0%BD%D1%8B%D0%B5_%D0%B3%D0%BB%D0%B0%D0%B7%D0%B0
- http://slovar.cc/biol/enc/2328012.html
- http://dic.academic.ru/dic.nsf/ruwiki/347188
- http://traditio.wiki/%D0%A4%D0%B0%D1%81%D0%B5%D1%82%D0%BE%D1%87%D0%BD%D1%8B%D0%B5_%D0%B3%D0%BB%D0%B0%D0%B7%D0%B0
- http://pest-nay.ru/mol/facetted-eyes-faceted-eyes-complex-eyes-the-main-paired-organ-of-sight-of-insects-crustaceans-and-some-other-invertebrates-are-formed-by-special-structural-units-ommatidia-afctnjxyst-ukfpf-encyclopedia-biology-ecology-animals-plant/