Кортикальные зрительные центры расположены на медиальной поверхности затылочной доли мозга в области шпорной борозды. Верхнюю губу шпорной борозды составляет cuneus, нижнюю — gyrus lingualis. Кора затылочной доли в области сиneus, gyrus lingualis и в глубине шпорной борозды представляет собою ядро кортикального конца зрительного анализатора. Последнее распространяется также и на наружную поверхность полюса затылочной доли, в той ее небольшой части, куда заходит задний конец шпорной борозды. Локализация зрительных центров в коре затылочной доли была установлена путем физиологических опытов на животных и клинико-анатомических исследований у человека.
В начале развития учения о высших зрительных центрах существовало весьма значительное расхождение между физиологическими исследованиями и данными клиники и патологической анатомии. Мунк впервые в опытах с удалением отдельных участков больших полушарий показал, что у собаки зрительный центр локализуется в затылочной доле. Он полагал, что центр этот расположен в определенной зоне наружной поверхности затылочной доли.
Бехтерев обратил внимание на то, что это воззрение, разделявшееся и другими авторами, стоит в резком противоречии с клиническими наблюдениями, согласно которым у человека зрительный центр расположен на медиальной поверхности затылочной доли. В опытах на собаках Бехтерев показал, что только разрушение медиальной поверхности затылочной доли приводит к стойким изменениям зрения по типу гемианопсии.
На основании своих опытов он пришел к выводу, что у собаки зрительный центр расположен на внутренней, а не на наружной поверхности затылочной доли. Тем самым были устранены расхождения, которые до этого имелись между экспериментальными данными и клиническими наблюдениями у человека. В дальнейшем Е. П. Кононова в опытах па собаках также подтвердила, что зрительный центр находится на медиальной поверхности затылочной доли.
Это было обосновано тремя группами фактических данных: 1) изменения в коре на наружной и нижней поверхности затылочной доли не сопровождаются развитием гемианопсии; 2) изменения коры медиальной поверхности затылочной доли в том случае, если они распространяются и на кору шпорной борозды, всегда приводят к гемианопсии; 3) гемианопсия наблюдается и тогда, когда изменения ограничиваются только корой области шпорной борозды.
Дальнейшие цитоархитектонические исследования показали, что кортикальные зрительные центры расположены в поле 17. Вместе с тем, однако, оставалось неясным, полностью ли кортикальные зрительные центры совпадают с распространением этого поля.
Полное совпадение кортикальных зрительных центров с полем 17 было установлено Г. Ленцем. Он исследовал кору затылочной доли при длительной, давностью от 8 до 39 лет, двусторонней слепоте. Во всех случаях он отмечал атрофию коры в области шпорной борозды. Как показали его цитоархитектонические исследования, атрофия занимала всю область восьмистопной коры (поле 17) и резко обрывалась у перехода в поля 18 и 19 с шестислойпой корон.
Вопрос о проекции сетчатки в области кортикальных зрительных центров является одним из наиболее сложных во всем учении о зрительных путях. Монаков и его последователи упорно отрицали существование такой проекции. По Монакову, нет постоянной связи отдельных участков сетчатки с определенными участками коры затылочной доли. Проекция. сетчатки на кору имеется лишь постольку, поскольку с детства проведение импульсов от отдельных участков сетчатки осуществляется избирательно вдоль определенных проводников и заканчивается в определенных участках коры.
Если эти участки разрушаются, то их функции замещаются новыми. Монаков полагал, что желтое пятно связано не с каким-нибудь ограниченным участком коры, а диффузно со всей корой зрительной области в целом.
Ошибочность этого воззрения была в дальнейшем доказана многолетними исследованиями Вильбранда и Геншена, отстаивавшими наличие строго определенной проекции сетчатки в коре затылочной доли.
Кора головного мозга — высший отдел центральной нервной системы, обеспечивающий функционирование организма как единого целого при его взаимодействии с окружающей средой.
Кора больших полушарий головного мозга (кора большого мозга, новая кора) представляет собой слой серого вещества, состоящего из 10-20 млрд нейронов и покрывающего большие полушария (рис. 1). Серое вещество коры составляет более половины всего серого вещества ЦНС. Суммарная площадь серого вещества коры — около 0,2 м 2 , что достигается извилистой складчатостью ее поверхности и наличием борозд разной глубины. Толщина коры в ее разных участках колеблется от 1,3 до 4,5 мм (в передней центральной извилине). Нейроны коры располагаются в шести слоях, ориентированных параллельно ее поверхности.
В участках коры, относящихся к лимбической системе, имеются зоны с трехслойным и пятислойным расположением нейронов в структуре серого вещества. Эти участки филогенетически древней коры занимают около 10% поверхности полушарий мозга, остальные 90% составляют новую кору.
Рис. 1. Моля латеральной поверхности коры большого мозга (по Бродману)
Строение коры головного мозга
Кора большого мозга имеет шестислойное строение
Нейроны разных слоев различаются по цитологическим признакам и функциональным свойствам.
Молекулярный слой — самый поверхностный. Представлен небольшим числом нейронов и многочисленными ветвящимися дендритами пирамидных нейронов, лежащих в более глубоких слоях.
Наружный зернистый слой сформирован плотно расположенными многочисленными мелкими нейронами разной формы. Отростки клеток этого слоя образуют кортикокортикальные связи.
Наружный пирамидальный слой состоит из пирамидных нейронов средней величины, отростки которых также участвуют в образовании кортикокортикальных связей между соседними областями коры.
Внутренний зернистый слой подобен второму слою по виду клеток и расположению волокон. В слое проходят пучки волокон, связывающие различные участки коры.
К нейронам этого слоя проводятся сигналы от специфических ядер таламуса. Слой очень хорошо представлен в сенсорных областях коры.
Внутренний пирамидный слои образован средними и крупными пирамидными нейронами. В двигательной области коры эти нейроны особенно крупные (50-100 мкм) и получили название гигантских, пирамидных клеток Беца. Аксоны этих клеток формируют быстропроводящие (до 120 м/с) волокна пирамидного тракта.
Слой полиморфных клеток представлен преимущественно клетками, аксоны которых образуют кортикоталамические пути.
Нейроны 2-го и 4-го слоев коры участвуют в восприятии, переработке поступающих к ним сигналов от нейронов ассоциативных областей коры. Сенсорные сигналы из переключающих ядер таламуса поступают преимущественно к нейронам 4-го слоя, выраженность которого наибольшая в первичных сенсорных областях коры. К нейронам 1-го и других слоев коры поступают сигналы из других ядер таламуса, базальных ганглиев, ствола мозга. Нейроны 3-го, 5-го и 6-го слоев формируют эфферентные сигналы, посылаемые в другие области коры и по нисходящим путям в нижележащие отделы ЦНС. В частности, нейроны 6-го слоя формируют волокна, следующие в таламус.
В нейронном составе и цитологических особенностях разных участков коры имеются значительные отличия. По этим отличиям Бродман разделил кору на 53 цитоархитектонических поля (см. рис. 1).
Расположение многих из этих нолей, выделенных на основе гистологических данных, совпадает по топографии с расположением корковых центров, выделенных на основе выполняемых ими функций. Используются и другие подходы деления коры на области, например, на основе содержания в нейронах определенных маркеров, по характеру нейронной активности и другим критериям.
Белое вещество полушарий головного мозга образовано нервными волокнами. Выделяют ассоциативные волокна, подразделяемые на дугообразные волокна, но которым сигналы передаются между нейронами рядом лежащих извилин и длинные продольные пучки волокон, доставляющие сигналы к нейронам более удаленных участков одноименного полушария.
Комиссуральные волокна — поперечные волокна, передающие сигналы между нейронами левого и правого полушарий.
Проекционные волокна — проводят сигналы между нейронами коры и других отделов мозга.
Перечисленные виды волокон участвуют в создании нейронных цепей и сетей, нейроны которых расположены на значительных расстояниях друг от друга. В коре имеется также особый вид локальных нейронных цепей, образованных рядом расположенными нейронами. Эти нейронные структуры получили название функциональных кортикальных колонок. Нейронные колонки образованы группами нейронов, расположенных друг над другом перпендикулярно поверхности коры. Принадлежность нейронов к одной и той же колонке можно определить по повышению их электрической активности на раздражение одного и того же рецептивного поля. Такая активность регистрируется при медленном перемещении регистрирующего электрода в коре в перпендикулярном направлении. Если регистрировать электрическую активность нейронов, расположенных в горизонтальной плоскости коры, то отмечается повышение их активности при раздражении различных рецептивных полей.
Диаметр функциональной колонки составляет до 1 мм. К нейронам одной функциональной колонки поступают сигналы от одного и того же афферентного таламокортикального волокна. Нейроны соседних колонок связаны друг с другом отростками, с помощью которых обмениваются информацией. Наличие в коре таких взаимосвязанных функциональных колонок увеличивает надежность восприятия и анализа информации, поступающей к коре.
Эффективность восприятия, обработки и использования информации корой для регуляции физиологических процессов обеспечивается также соматотопическим принципом организации сенсорных и моторных полей коры. Суть такой организации заключается в том, что в определенной (проекционной) области коры представлены не любые, а топографически очерченные участки рецептивного поля поверхности тела, мышц, суставов или внутренних органов. Так, например, в соматосенсорной коре поверхность тела человека спроецирована в виде схемы, когда в определенной точке коры представлены рецептивные поля конкретной области поверхности тела. Строгим топографическим образом в первичной моторной коре представлены эфферентные нейроны, активация которых вызывает сокращение определенных мышц тела.
Полям коры присущ также экранный принцип функционирования. При этом рецепторный нейрон посылает сигнал не на одиночный нейрон или в одиночную точку коркового центра, а на сеть или ноле нейронов, связанных отростками. Функциональными ячейками этого поля (экрана) являются колонки нейронов.
Кора мозга, формируясь на поздних этапах эволюционного развития высших организмов, в определенной мере подчинила себе все нижележащие отделы ЦНС и способна корригировать их функции. В то же время функциональная активность коры больших полушарий определяется притоком к ней сигналов от нейронов ретикулярной формации ствола мозга и сигналов от рецептивных полей сенсорных систем организма.
Функциональные области коры мозга
По функциональному признаку в коре выделяют сенсорные, ассоциативные и двигательные области.
Сенсорные (чувствительные, проекционные) области коры
Они состоят из зон, содержащих нейроны, активация которых афферентными импульсами от сенсорных рецепторов или прямым воздействием раздражителей вызывает появление специфических ощущений. Эти зоны имеются в затылочной (поля 17-19), теменной (ноля 1-3) и височной (поля 21-22, 41-42) областях коры.
В сенсорных зонах коры выделяют центральные проекционные поля, обеспечивающие топкое, четкое восприятие ощущений определенных модальностей (свет, звук, прикосновение, тепло, холод) и вторичные проекционные ноля. Функцией последних является обеспечение понимания связи первичного ощущения с другими предметами и явлениями окружающего мира.
Зоны представительства рецептивных полей в сенсорных зонах коры в значительной мере перекрываются. Особенность нервных центров в области вторичных проекционных полей коры — их пластичность, которая проявляется возможностью перестройки специализации и восстановления функций после повреждения какого-либо из центров. Эти компенсаторные возможности нервных центров особенно выражены в детском возрасте. В то же время повреждение центральных проекционных полей после перенесенных заболевании, сопровождается грубым нарушением функций чувствительности и часто невозможностью ее восстановления.
Зрительная кора
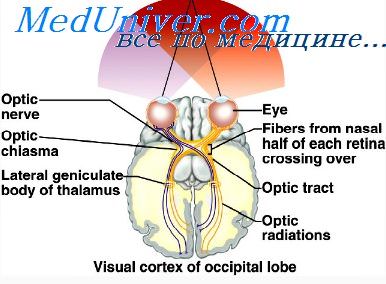
Первичная зрительная кора (VI, поле 17) располагается по обеим сторонам шпорной борозды на медиальной поверхности затылочной доли головного мозга. В соответствии с выявлением па неокрашенных срезах зрительной коры чередующихся белых и темных полос ее называют также стриарной (полосатой) корой. К нейронам первичной зрительной коры посылают зрительные сигналы нейроны латерального коленчатого тела, которые получают сигналы от ганглиозных клеток сетчатки. Зрительная кора каждого полушария получает визуальные сигналы от ипсилатеральной и контралатеральной половин сетчатки обоих глаз и их поступление к нейронам коры организовано по соматотопическому принципу. Нейроны, к которым поступают зрительные сигналы от фоторецепторов, топографически расположены в зрительной коре подобно рецепторам в сетчатке глаза. При этом область желтого пятна сетчатки имеет относительно большую зону представительства в коре, чем другие области сетчатки.
Нейроны первичной зрительной коры ответственны за зрительное восприятие, которое на основе анализа входных сигналов проявляется их способностью обнаруживать зрительный стимул, определять его специфическую форму и ориентацию в пространстве. Упрощенно можно представить сенсорную функцию зрительной коры в решении задачи и ответе на вопрос, что представляет собой зрительный объект.
В анализе других качеств зрительных сигналов (например, расположения в пространстве, движения, связи с другими событиями и т.д.) принимают участие нейроны полей 18 и 19 экстрастриарной коры, расположенных но соседству с нолем 17. Информация о сигналах, поступивших в сенсорные зрительные зоны коры, передастся для дальнейшего анализа и использования зрения для выполнения других функций мозга в ассоциативные области коры и другие отделы мозга.
Слуховая кора
Расположена в латеральной борозде височной доли в области извилины Гешля (AI, поля 41-42). К нейронам первичной слуховой коры поступают сигналы от нейронов медиальных коленчатых тел. Волокна слуховых путей, проводящие звуковые сигналы в слуховую кору, организованы тонотопически, и это позволяет нейронам коры получать сигналы от определенных слуховых рецепторных клеток кортиева органа. Слуховая кора регулирует чувствительность слуховых клеток.
В первичной слуховой коре формируются звуковые ощущения и проводится анализ отдельных качеств звуков, позволяющий ответить на вопрос, что представляет собой воспринятый звук. Первичная слуховая кора играет важную роль в анализе коротких звуков, интервалов между звуковыми сигналами, ритма, звуковой последовательности. Более сложный анализ звуков осуществляется в ассоциативных областях коры, смежных с первичной слуховой. На основе взаимодействия нейронов этих областей коры осуществляется бинауральный слух, определяются характеристики высоты, тембра, громкости звука, принадлежность звука, формируется представление о трехмерном звуковом пространстве.
Вестибулярная кора
Располагается в верхней и средней височных извилинах (поля 21-22). К ее нейронам поступают сигналы от нейронов вестибулярных ядер ствола мозга, связанных афферентными связями с рецепторами полукружных каналов вестибулярного аппарата. В вестибулярной коре формируется ощущение о положении тела в пространстве и ускорении движений. Вестибулярная кора взаимодействует с мозжечком (через височно-мостомозжечковый путь), участвует в регуляции равновесия тела, приспособлении позы к осуществлению целенаправленных движений. На основе взаимодействия этой области с соматосенсорной и ассоциативными областями коры происходит осознание схемы тела.
Обонятельная кора
Расположена в области верхней части височной доли (крючок, ноля 34, 28). Кора включает ряд ядер и относится к структурам лимбической системы. Ее нейроны расположены в трех слоях и получают афферентные сигналы от митральных клеток обонятельной луковицы, связанных афферентными связям с обонятельными рецепторными нейронами. В обонятельной коре проводится первичный качественный анализ запахов и формируется субъективное ощущение запаха, его интенсивности, принадлежности. Повреждение коры ведет к снижению обоняния или к развитию аносмии — потере обоняния. При искусственном раздражении этой области возникают ощущения различных запахов по типу галлюцинаций.
Вкусовая кора
Расположена в нижней части соматосенсорной извилины, непосредственно кпереди от области проекции лица (поле 43). Ее нейроны получают афферентные сигналы от релейных нейронов таламуса, которые связаны с нейронами ядра одиночного тракта продолговатого мозга. К нейронам этого ядра поступают сигналы непосредственно от чувствительных нейронов, образующих синапсы на клетках вкусовых луковиц. Во вкусовой коре проводится первичный анализ вкусовых качеств горького, соленого, кислого, сладкого и на основе их суммации формируется субъективное ощущение вкуса, его интенсивности, принадлежности.
Сигналы запахов и вкуса достигают нейронов передней части островковой коры, где на основе их интеграции формируется новое, более сложное качество ощущений, определяющее наше отношение к источникам запаха или вкуса (например, к пище).
Соматосенсорная кора
Занимает область постцентральной извилины (SI, поля 1-3), включая парацентральную дольку на медиальной стороне полушарий (рис. 9.14). В соматосенсорную область поступают сенсорные сигналы от нейронов таламуса, связанных спиноталамическими путями с рецепторами кожи (тактильная, температурная, болевая чувствительность), проприорецепторами (мышечных веретен, суставных сумок, сухожилий) и интерорецепторами (внутренних органов).
Рис. 9.14. Важнейшие центры и области коры большого мозга
Из-за перекреста афферентных путей в соматосенсорную зону левого полушария приходит сигнализация от правой стороны тела, соответственно в правое полушарие — от левой стороны тела. В этой сенсорной области коры соматотопически представлены все части тела, но при этом наиболее важные рецептивные зоны пальцев рук, губ, кожи лица, языка, гортани занимают относительно большие площади, чем проекции таких поверхностей тела, как спина, передняя часть туловища, ноги.
Расположение представительства чувствительности частей тела вдоль постцентральной извилины часто называют «перевернутый гомункулюс», так как проекция головы и шеи находится в нижней части постцентральной извилины, а проекция каудальной части туловища и ног — в верхней части. При этом чувствительность голеней и стоп проецируется на кору пара- центральной дольки медиальной поверхности полушарий. Внутри первичной соматосенсорной коры имеется определенная специализация нейронов. Например, нейроны поля 3 получают преимущественно сигналы от мышечных веретен и механорецепторов кожи, поля 2 — от рецепторов суставов.
Кору постцентральной извилины относят к первичной соматосенсорной области (SI). Ее нейроны посылают обработанные сигналы к нейронам вторичной соматосенсорной коры (SII). Она располагается кзади от постцентральной извилины в теменной коре (поля 5 и 7) и принадлежит к ассоциативной коре. Нейроны SII не получают прямых афферентных сигналов от нейронов таламуса. Они связаны с нейронами SI и нейронами других областей коры мозга. Это позволяет проводить здесь интегральную оценку сигналов, попадающих в кору по спиноталамическому пути с сигналами, поступающими из других (зрительной, слуховой, вестибулярной и т.д.) сенсорных систем. Важнейшей функцией этих полей теменной коры является восприятие пространства и трансформация сенсорных сигналов в координаты моторных. В теменной коре формируется стремление (намерение, побуждение) осуществить моторное действие, что является основой для начала планирования в ней предстоящей моторной активности.
Интеграция различных сенсорных сигналов связана с формированием различных ощущений, адресуемых к разным частям тела. Эти ощущения используются как для формирования психических, так и других ответных реакций, примерами которых могут быть движения при одновременном участии мышц обеих сторон тела (например, перемещение, ощупывание обеими руками, хватание, однонаправленное движение обеими руками). Функционирование этой области необходимо для узнавания предметов на ощупь и определения пространственного расположения этих предметов.
Нормальная функция соматосенсорных областей коры является важным условием формирования таких ощущений как тепло, холод, боль и их адресации к определенной части тела.
Повреждение нейронов области первичной соматосенсорной коры ведет к снижению различных видов чувствительности на противоположной стороне тела, а локальное повреждение — к потере чувствительности в определенной части тела. Особенно ранимой при повреждении нейронов первичной соматосенсорной коры является дискриминационная чувствительность кожи, а наименее — болевая. Повреждение нейронов вторичной соматосенсорной области коры может сопровождаться нарушением способности распознания предметов на ощупь (тактильная агнозия) и навыков использования предметов (апраксия).
Двигательные области коры
Около 130 лет тому назад исследователи, нанося точечные раздражения на кору мозга электрическим током, обнаружили, что воздействие на поверхность передней центральной извилины вызывает сокращение мышц противоположной стороны тела. Так было обнаружено наличие одной из моторных зон коры мозга. В последующем оказалось, что к организации движений имеют отношение несколько областей коры мозга и его другие структуры, а в областях моторной коры имеются не только двигательные нейроны, но и нейроны, осуществляющие другие функции.
Первичная моторная кора
Первичная моторная кора располагается в передней центральной извилине (MI, поле 4). Ее нейроны получают основные афферентные сигналы от нейронов соматосенсорной коры — полей 1, 2, 5, премоторной коры и таламуса. Кроме того, через вентролатеральный таламус в MI посылают сигналы нейроны мозжечка.
От пирамидных нейронов Ml начинаются эфферентные волокна пирамидного пути. Часть волокон этого пути следует к моторным нейронам ядер черепных нервов ствола мозга (кортикобульбарный тракт), часть — к нейронам стволовых моторных ядер (красное ядро, ядра ретикулярной формации, стволовые ядра, связанные с мозжечком) и часть — к интер- и моторным нейронам спинного мозга (кортикоспинальный тракт).
Имеется соматотопическая организация расположения нейронов в MI, контролирующих сокращение разных мышечных групп тела. Нейроны, контролирующие мышцы ног и туловища, расположены в верхних участках извилины и занимают относительно малую площадь, а контролирующие мышцы рук, особенно пальцев, лица, языка и глотки расположены в нижних участках и занимают большую площадь. Таким образом, в первичной двигательной коре относительно большую площадь занимают те нейронные группы, которые управляют мышцами, осуществляющими разнообразные, точные, мелкие, тонко регулируемые движения.
Поскольку многие нейроны Ml увеличивают электрическую активность непосредственно перед началом произвольных сокращений, то первичной моторной коре отводят ведущую роль в контроле активности моторных ядер ствола и мотонейронов спинного мозга и инициации произвольных, целенаправленных движений. Повреждение поля Ml ведет к парезу мышц и невозможности осуществления тонких произвольных движений.
Вторичная моторная кора
Включает области премоторной и дополнительной моторной коры (МII, поле 6). Премоторная кора расположена в поле 6, на боковой поверхности мозга, кпереди от первичной моторной коры. Ее нейроны получают через таламус афферентные сигналы из затылочной, соматосенсорной, теменной ассоциативной, префронтальной областей коры и мозжечка. Обработанные в ней сигналы нейроны коры посылают по эфферентным волокнам в моторную кору MI, небольшое число — в спинной мозг и большее — в красные ядра, ядра ретикулярной формации, базальные ганглии и мозжечок. Премоторная кора играет основную роль в программировании и организации движений, находящихся под контролем зрения. Кора участвует в организации позы и вспомогательных движений для действий, осуществляемых дистальными мышцами конечностей. Повреждение прсмотор- ной коры часто вызывает тенденцию повторного выполнения начатого движения (персеверация), даже если осуществленное движение достигло цели.
В нижней части премоторной коры левой лобной доли, непосредственно кпереди от участка первичной моторной коры, в которой представлены нейроны, контролирующие мышцы лица, располагается речевая область, или моторный центр речи Брока. Нарушение ее функции сопровождается нарушением артикуляции речи, или моторной афазией.
Дополнительная моторная кора располагается в верхней части поля 6. Ее нейроны получают афферентные сигналы из соматосснсорной, теменной и префронтальной областей коры головного мозга. Обработанные в ней сигналы нейроны коры посылают по эфферентным волокнам в первичную моторную кору MI, спинной мозг, стволовые моторные ядра. Активность нейронов дополнительной моторной коры повышается раньше, чем нейронов коры MIи главным образом в связи с осуществлением сложных движений. При этом возрастание нейронной активности в дополнительной моторной коре не связано с движениями как таковыми, для этого достаточно мысленно представить модель предстоящих сложных движений. Дополнительная моторная кора принимает участие в формировании программы предстоящих сложных движений и в организации моторных реакций на специфичность сенсорных стимулов.
Поскольку нейроны вторичной моторной коры посылают множество аксонов в поле MI, ее считают в иерархии моторных центров организации движений более высокой структурой, стоящей над моторными центрами моторной коры MI. Нервные центры вторичной моторной коры могут оказывать влияние на активность моторных нейронов спинного мозга двумя путями: непосредственно через кортикоспинальный путь и через поле MI. Поэтому их иногда называют супрамоторными полями, в функцию которых входит инструктирование центров поля MI.
Из клинических наблюдений известно, что сохранение нормальной функции вторичной моторной коры важно для осуществления точных движений руки, и особенно для выполнения ритмических движений. Так, например, при их повреждении пианист перестает чувствовать ритм и выдерживать интервал. Нарушается способность к осуществлению противоположных движений руками (манипулирование обоими руками).
При одновременном повреждении моторных зон MI и MII коры утрачивается способность к тонким координированным движениям. Точечные раздражения в этих областях моторной зоны сопровождаются активацией не отдельных мышц, а целой группы мышц, вызывающих направленное движение в суставах. Эти наблюдения послужили поводом для формирования вывода о том, что в моторной коре представлены не столько мышцы, сколько движения.
Располагается в области поля 8. Ее нейроны получают основные афферентные сигналы из затылочной зрительной, теменной ассоциативной коры, верхних холмиков четверохолмия. Обработанные сигналы передаются по эфферентным волокнам в премоторную кору, верхние холмики четверохолмия, стволовые моторные центры. Кора играет определяющую роль в организации движений, находящихся под контролем зрения и принимает непосредственное участие в инициации и контроле движений глаз и головы.
Механизмы, реализующие превращение замысла движения в конкретную моторную программу, в залпы импульсов, посылаемых к определенным мышечным группам, остаются недостаточно понятными. Считается, что замысел движения формируется благодаря функциям ассоциативной и других областей коры, взаимодействующих со многими структурами головного мозга.
Информация о замысле движения передается в двигательные области лобной коры. Двигательная кора через нисходящие пути активирует системы, обеспечивающие выработку и использование новых двигательных программ или использование старых, уже отработанных на практике и хранящихся в памяти. Составной частью этих систем являются базальные ганглии и мозжечок (см. их функции выше). Программы движения, выработанные при участии мозжечка и базальных ганглиев, передаются через таламус в моторные зоны и прежде всего в первичную моторную область коры. Эта область непосредственно инициирует исполнение движений, подключая к нему определенные мышцы и обеспечивая последовательность смены их сокращения и расслабления. Команды коры передаются на моторные центры ствола мозга, спинальные мотонейроны и мотонейроны ядер черепных нервов. Мотонейроны в осуществлении движений выполняют роль конечного пути, через который двигательные команды передаются непосредственно к мышцам. Особенности передачи сигналов от коры к моторным центрам ствола и спинного мозга описаны в главе, посвященной ЦНС (ствол мозга, спинной мозг).
Ассоциативные области коры
У человека ассоциативные области коры занимают около 50% площади всей коры большого мозга. Они располагаются в участках между сенсорными и двигательными областями коры. Ассоциативные области не имеют четких границ со вторичными сенсорными областями как по морфологическим, так и по функциональным признакам. Выделяют теменную, височную и лобную ассоциативные области коры больших полушарий.
Теменная ассоциативная область коры. Располагается в полях 5 и 7 верхней и нижней теменных долек мозга. Область граничит впереди с соматосенсорной корой, сзади — со зрительной и слуховой корой. К нейронам теменной ассоциативной области могут поступать и активировать их зрительные, звуковые, тактильные, проприоцептивные, болевые, сигналы из аппарата памяти и другие сигналы. Часть нейронов является полисенсорной и может повышать свою активность при поступлении к ней соматосенсорных и визуальных сигналов. Однако степень повышения активности нейронов ассоциативной коры на поступление афферентных сигналов зависит от текущей мотивации, внимания субъекта и информации, извлекаемой из памяти. Она остается незначительной, если поступающий из сенсорных областей мозга сигнал для субъекта безразличен, и существенно возрастает, если он совпал с имеющейся мотивацией и привлек его внимание. Например, при предъявлении обезьяне банана активность нейронов ассоциативной теменной коры остается невысокой, если животное сыто, и наоборот, активность резко возрастает у голодных животных, которым нравятся бананы.
Нейроны теменной ассоциативной коры связаны эфферентными связями с нейронами префронтальной, премоторной, моторной областей лобной доли и поясной извилины. Исходя из экспериментальных и клинических наблюдений, принято считать, что одной из функций коры поля 5 является использование соматосенсорной информации для осуществления целенаправленных произвольных движений и манипулирования объектами. Функцией коры поля 7 является интеграция визуальных и соматосенсорных сигналов для координации движений глаз и визуально-ведомых движений руки.
Нарушением этих функций теменной ассоциативной коры при повреждении ее связей с корой лобной доли или заболеванием самой лобной доли, объясняются симптомы последствий заболеваний, локализованных в области теменной ассоциативной коры. Они могут проявляться затруднением в понимании смыслового содержания сигналов (агнозия), примером которого может быть потеря способности распознавания формы и пространственного расположения объекта. Могут нарушаться процессы трансформации сенсорных сигналов в адекватные моторные действия. В последнем случае больной теряет навыки практического использования хорошо знакомых инструментов и предметов (апраксия), и у него может развиться невозможность осуществления визуально-ведомых движений (например, движение руки в направлении предмета).
Лобная ассоциативная область коры. Располагается в префронтальной коре, которая является частью коры лобной доли, локализующейся кпереди от полей 6 и 8. Нейроны лобной ассоциативной коры получают обработанные сенсорные сигналы по афферентным связям от нейронов коры затылочной, теменной, височной долей мозга и от нейронов поясной извилины. Лобная ассоциативная кора получает сигналы о текущем мотивационном и эмоциональном состояниях от ядер таламуса, лимбической и других структур мозга. Кроме того, лобная кора может оперировать абстрактными, виртуальными сигналами. Эфферентные сигналы ассоциативная лобная кора посылает обратно, в структуры мозга, от которых они были получены, в моторные области лобной коры, хвостатое ядро базальных ганглиев и гипоталамус.
Эта область коры играет первостепенную роль в формировании высших психических функций человека. Она обеспечивает формирование целевых установок и программ осознанных поведенческих реакций, узнавание и смысловую оценку предметов и явлений, понимание речи, логическое мышление. После обширных повреждений лобной коры у больных могут развиться апатия, снижение эмоционального фона, критичного отношения к своим собственным поступкам и поступкам окружающих, самодовольство, нарушение возможности использования прошлого опыта для изменения поведения. Поведение больных может стать непредсказуемым и неадекватным.
Височная ассоциативная область коры. Располагается в полях 20, 21, 22. Нейроны коры получают сенсорные сигналы от нейронов слуховой, экстрастриарной зрительной и префронтальной коры, гиппокампа и миндалины.
После двухстороннего заболевания височных ассоциативных областей с вовлечением в патологический процесс гиппокампа или связей с ним у больных могут развиться выраженные нарушения памяти, эмоционального поведения, неспособность сосредоточения внимания (рассеянность). У части людей при повреждении нижневисочной области, где предположительно располагается центр узнавания лица, может развиться зрительная агнозия — неспособность узнавания лиц знакомых людей, предметов, при сохранности зрения.
На границе височной, зрительной и теменной областей коры в нижней теменной и задней части височной доли располагается ассоциативный участок коры, получивший название сенсорного центра речи, или центра Вернике. После его повреждения развивается нарушение функции понимания речи при сохранности речедвигательной функции.
Лекция 4 Корковые центры коры больших полушарий головного мозга.
Понятие корковый центр предложено И.П.Павловым. Корковый центр не имеет четких границ и состоит из ядерных и рассеянных частей. Корковые центры располагаются в долях головного мозга.
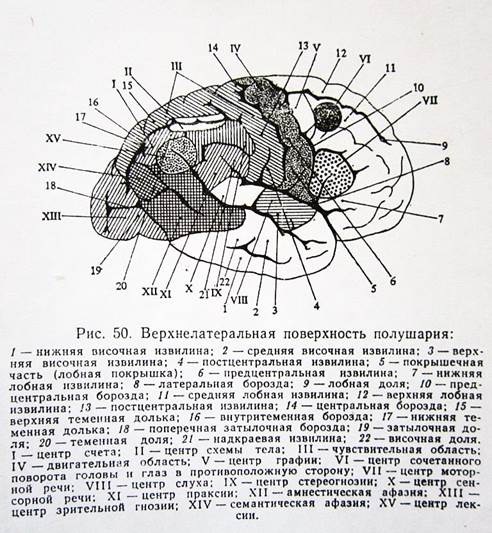
I. Лобная доля головного мозга.
1. Двигательная зона расположена в предцентральной извилине. В верхней трети располагаются нейроны, иннервирующие ногу, в средней – руку, в нижней – лицо, язык, гортань и глотку. Раздражение слабым электрическим током этой зоны приводит к сокращению определенной группы мышц. При нарушениях двигательной зоны головного мозга возникают парезы (ослабление движений) и параличи (полное отсутствие движений).
2. Центр сочетанного поворота головы и глаз располагается в средней лобной извилине. Является двусторонним центром и осуществляет сочетанный поворот головы и глаз в противоположную сторону. При повреждении центра в правом полушарии, голова и глаза смотрят в правую сторону, т.е. в сторону повреждения. Больной с таким повреждением не может осуществлять поворот головы и глаз в противоположную поврежденной части сторону.
3. Центр моторной речи (центр Брока) – расположен в заднем нижнем отделе третьей лобной извилины левого полушария (у правшей). При повреждении нарушается моторная речь (способность говорить).
4. Центр письма (центр графии) располагается задних отделах средней лобной извилины. При повреждении нарушается способность писать (аграфия).
II. Теменная доля.
1. Центр общих видов чувствительности – располагается в теменной доле за центральной бороздой (борозда Роланда). Представляет собой двусторонний центр. В верхнем отделе расположены чувствительные центры ноги, в среднем отделе – руки, в нижнем – головы. При повреждении центра возникают нарушения чувствительности соответствующих органов.
2. Центр восприятия сложных видов чувствительности – двухсторонний центр, расположенный в верхней теменной дольке. Отвечает за восприятие веса, двухмерности и пр.
3. Центр схемы тела – располагается внутри теменной борозды. Поражения центра приводят к нарушениям правильных представлений о размерах, форме, расположении частей своего тела.
4. Центр праксии – расположен над краевой извилиной обоих полушарий. Обеспечивает выполнение сложных целенаправленных движений в определенной последовательности. При поражении центра возникают нарушения целенаправленных движений и действий при сохранности составляющих его элементарных движений (апраксии). Апраксии бывают трех видов:
— моторная (апраксия выполнения) – расстройство действий по приказу или подражанию;
— конструктивная – невозможность сконструировать цельное движение из отдельных его частей.
5. Центр стереогнозии – располагается позади средней части постцентральной извилины. При повреждении нарушается способность узнавать предметы наощупь.
6. Центр лексии – располагается в угловой извилине левого полушария у правши. Отвечает за способность узнавать печатные знаки и читать. При нарушениях центра возникает алексия – нарушение чтения.
7. Центр счета (калькулии) – располагается над угловой извилиной. При поражении центра нарушается способность считать – акалькулия.
8. Центр семантической афазии – расположен в области схода теменной, затылочной и височной долей. При повреждении нарушается способность понимать сложные словесно-грамматические, временные, смысловые конструкции.
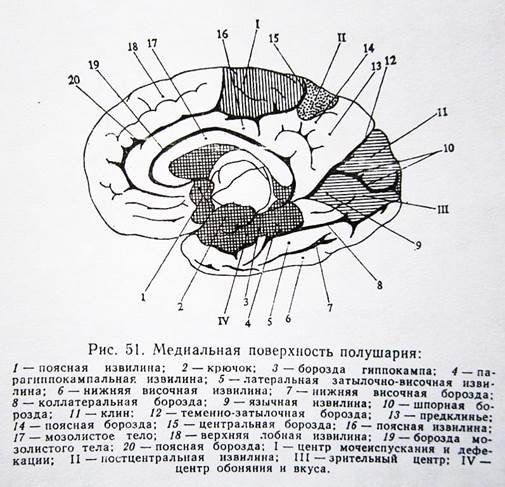
III. Височная доля.
1. Центр сенсорной речи (центр Вернике) – располагается в заднем отделе верхневисочной извилины левого полушария у правшей. Отвечает за понимание устной речи. Повреждение центра приводит к сенсорной афазии – нарушению понимания устной речи.
2. Центр амнестической афазии. При повреждении нарушается способность называть предметы, назначение которых хорошо известно больному.
3. Центр слуха – двухсторонний центр, располагается в верхневисочной извилине и частично в поперечной височной извилине. При повреждении нарушается слышимость на оба уха, но в большей степени на противоположное, т.е. при повреждении в левом полушарии больше страдает слышимость правого уха.
4. Центр вкуса и обоняния – располагается в области гиппокампа. При повреждении нарушается способность воспринимать вкус и запах.
IV. Затылочная доля.
1. Центр зрения – расположен в области шпорной борозды и клиновидной извилины. При повреждении возникают нарушения зрения, выпадения полей зрения.
2. Центр зрительной гнозии – расположен верхнелатеральной поверхности затылочной доли. При повреждении нарушается способность узнавать предметы.
Лекция 6 Мозжечок
Мозжечок является отделом ЦНС. Принимает участие в координации движений, регуляции равновесия, точности, соразмерности и правильности движений, регулирует мышечный тонус, является важнейшим центром вегетативной (?) нервной системы.
Располагается в задней черепной ямке позади ствола головного мозга. Состоит из полушарий мозжечка и червя мозжечка. Червь мозжечка отвечает за статическую координацию, полушария мозжечка – за динамическую координацию движений. Соматотопически в черве мозжечка представлены мышцы туловища, в полушариях – мышцы конечностей.
Поверхность мозжечка покрыта слоем серого вещества, составляющим кору мозжечка. Кора мозжечка покрыта тонкими бороздами и извилинами. Есть несколько долей мозжечка.
Белое вещество образует ножки мозжечка, в которых расположены проводящие пути. Ножки мозжечка делятся на нижние, средние и верхние. Нижние ножки соединены с продолговатым мозгом, средние – с Варолиевым мостом, верхние – со средним мозгом.
В белом веществе мозжечка имеются ядра серого вещества:
2. Шаровидное ядро;
3. Зубчатое ядро;
4. Пробковидное ядро.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Химия, Биология, подготовка к ГИА и ЕГЭ
«… мозг есть орган души, т. е. такой механизм, который будучи приведен какими ни на есть причинами в движение, дает в окончательном результате тот ряд внешних явлений, которыми характеризуется психическая деятельность».
автор статьи — Мария Щербакова
Мозг человека – сложнейший по своей организации и совершенный, по сути, орган.
Подумать только, он обеспечивает все в нашей жизни: возможность ходить, дышать, видеть, слышать, говорить, думать, жить!
Мозг координирует и регулирует все жизненно важные функции организма человека, более того мозг контролирует его поведение.
Если мозг перестает работать, то организм человека переходит в пассивное состояние, когда на любую стимуляцию, извне или изнутри, нет реакции. Человек не сможет слышать, видеть, чувствовать, осознанно двигаться – он как овощ, который просто существует, но в полной изоляции, депривации от внешнего мира.
Все мы знаем, что мозг высшего млекопитающего делится на две основные части: спинной и головной.
Головной мозг по своей структуре симметричен.
При рождении малыша его мозг весит примерно 300 г,
по мере роста человека он увеличивается и у взрослого он весит около 1500 г.
мозг мужчин, как правило, чуть тяжелее мозга женщины.
У здорового взрослого индивида вес мозга составляет около 2% от общего веса человека.
Не стоит думать, что чем больше весит мозг, тем умнее и гениальнее человек. Ученые давно доказали, что уровень интеллекта и гениальность совершенно не связаны с весом мозга.
Гениальность и интеллект зависят от количества нервных связей, которые создает сам мозг.
Что же представляет собой головной мозг человека, какие отделы он содержит?
1) Продолговатый мозг, который управляет вегетативными функциями организма человека.
Он отвечает, прежде всего, за регуляцию дыхания, сердечно-сосудистую деятельность, пищеварительные рефлексы, обмен веществ.
2) Задний мозг: мозжечок и варолиев мост.
Именно он ответственен, за координацию движений
3) Средний мозг — отвечает за первичные ориентировочные рефлексы организма человека на внешние раздражители.
Движение глаз, поворот головы в сторону источника звука или света — это работа среднего мозга, так называемого нашего зрительного центра.
4) Промежуточный мозг:
а) таламус, который обеспечивает обработку большей части импульсов от наших рецепторов (ну кроме обонятельных), а также отвечает за эмоциональную окраску информации;
б) гипоталамус, который регулирует вегетативные функции организма
В нем расположены центры чувства насыщения, голода, жажды, удовольствия, и обеспечивает регуляцию сна и бодрствования человека.
5) Передний мозг представляет собой два полушария: левое и правое. Его поверхность покрыта бороздами и извилинами, что увеличивает поверхность, следовательно, обеспечивает более совершенную работу мозга. Полушария составляют до 80% массы всего мозга.
Благодаря коре больших полушарий возможна работа высших психических функций.
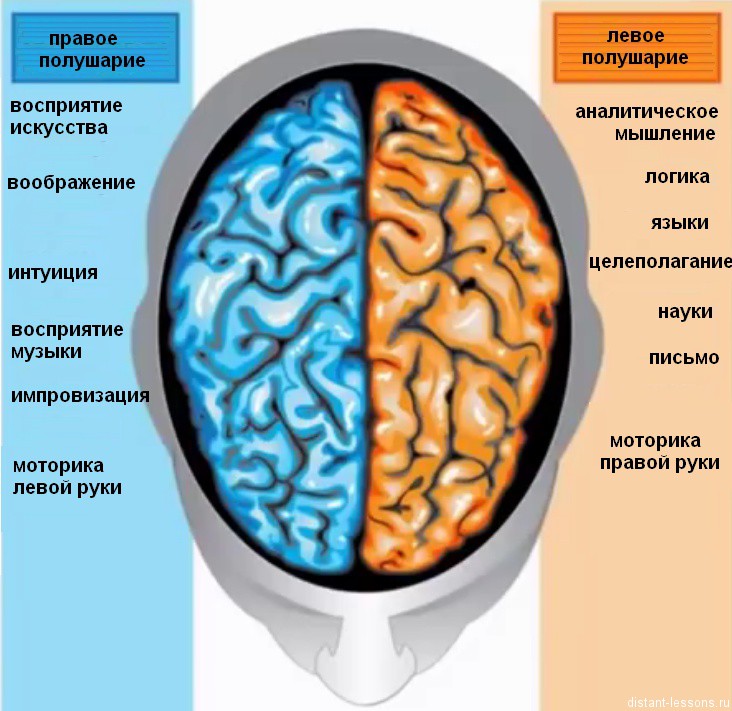
Считается, что левое полушарие отвечает за мыслительные процессы, счет и письмо, а правое – за восприятие сигналов внешнего мира. Левое полушарие – абстрактно-логическое, правое – творческое и образное.
Однако в настоящее время ученые считают такое деление достаточно условным, потому что в реализации высшей психической деятельности человека, его поведении участвуют равнозначно оба полушария, хотя конечно они играют различную роль в формировании образов восприятия.
Кора головного мозга отвечает за ряд специфических функций.
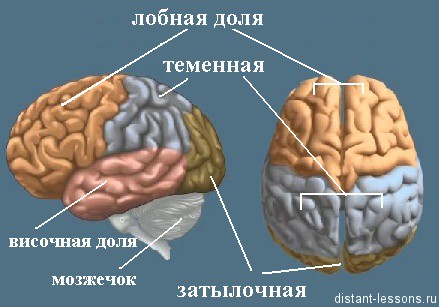
Височная доля отвечает за слух и обоняние,
затылочная за зрение,
теменная за осязание и вкус,
лобная за речь, движение и мышление.
Причем чем сложнее действие, тем большая часть коры отвечает за него.
В психологии и нейропсихологии существует такое понятие как гомункулус.
Гомункулус – это некая физиологическая и психологическая метафора.
Средневековые алхимики говорили о существе, подобном человеческому, которое можно создать искусственным путем. Например, Парацельс в XVI веке предлагал такой «рецепт»: человеческую сперму, необходимо заключить в особый сосуд, потом провести с ней длительные процессы обработки (некие манипуляции) и она станет гомункулом, который надо «вскармливать» человеческой кровью.
В XVII-XVIII веках считалось, что гомункул содержится в сперме человека, а когда попадает в организм будущей мамы, то превращается в человека. Гомункул выступает здесь как «ген передачи», некое существо, живущее в теле человека, регулирующее его мораль и ценности, который управляет поведением человека.
Конечно это лишь предположения и догадки свойственные развитию мысли и науки того времени. Однако термин остался и прижился для определения сложной работы коры головного мозга человека.
Выходит, гомункулус в современной науке – это схематическое изображение моторных и сенсорных функций человека на корковой проекции. Мы видим пропорции тела человека, его функций и действий, его поведение, в соотношении к количеству коры, задействованной в работе этих функций.
Чем сложнее действие, чем мельче моторика, чем выше психическая функция, тем большая площадь коры за него отвечает.
1) нормальная работа его отделов обеспечивает функционирование всего организма, здоровье человека, возможность деятельности человека, его потенциал, его реакцию на все виды раздражителей, его поведенческие реакции.
2) работа больших полушарий — функционирование коры головного мозга, которая обеспечивает весь широкий спектр его психических функций: ощущение и восприятие, внимание, мышление и речь, его память, воображение и т.д.– словом все то, что составляет сущность его психической деятельности, его сознания.
Сознание человека – это высшая форма отражения действительности, оно связано, самым тесным образом, с работой мозга человека: с речью, мышлением (абстрактным и логическим), памятью. Сознание – это функция мозга
Именно оно обеспечивает единство и регулирование человеческой деятельности и поведения.